被子植物

物种起源
概述
陆生植物已存在了约4.25亿年之久,从第一个以其水栖同类的简单方式-孢子传播生殖的植物之来。在海里,植物(和一些动物)能简单地将其自身的部份小复制体传播出去,让它们四处浮动和生长。人们认为这便是早期的植物(如现今的蕨类)的繁殖方式。但植物很快地便发展出保护其复制体,如干旱和其他在陆地上比在海洋里很合适的多余措施来。这类保护变成了种子,但还不是花。早期的种子植物包括有银杏和松科(如松属和冷杉属)。花粉-被认为和花朵的发展有最直接的关连,其被发现的化石最早则可上推至1.3亿年前。接近近代的花朵化石的突然出现,在演化论中产生了一个问题,被达尔文称做是“恼人之谜”(abominable mystery)。
已灭绝裸子植物的一些类型,尤其是种子蕨,被推测是开花植物的祖先,但之中并没有直接的连续化石证据来表示花朵是如何演化出来的。一些较早的化石,如三叠纪早期的“Sanmiguelia”即被推测是开花植物的祖先之一。依据如今的证据,一些人亦推测开花植物的祖先是由于三叠纪晚期(2.45亿至2.02亿年前)的未知裸子植物分歧出来的。早期的大羽羊齿类植物至开花植物间的关系仍然是个谜。
被子植物和买麻藤纲之间亲近的关系-依其形态上的证据,因分子上的分析而出现了争议,而使得买麻藤纲被认为更接近其他的裸子植物。
 现代的基因分析(分子系统发生)表示,在太平洋的岛屿-新喀里多尼亚上发现的互叶梅是其他开花植物的旁系,且形态上的研究也推测其可能有着最早开花植物特征的外表。
现代的基因分析(分子系统发生)表示,在太平洋的岛屿-新喀里多尼亚上发现的互叶梅是其他开花植物的旁系,且形态上的研究也推测其可能有着最早开花植物特征的外表。
巨大的开花植物适应辐射(当其在化石上出现著巨大分歧时)出如今白垩纪中期(约1亿年前)。而在白垩纪晚期时,开花植物已成了陆生植物的优势种族,且许多被认为可归至现今的科内的化石植物(包括山毛榉、栎树、枫和木兰等)也在此时出现。
许多花朵的颜色和形状,一般猜测,花朵的功能从一开始就将其他动物包含进了其生殖的过程中。花粉传播可以不需要鲜艳的颜色和明显的形状,因此这会是个累赘,平白地浪费了植物的养分,除非它们能提供其他的一些好处。对于花朵那突然且完整发展出来的外观,其中一个猜想是因为它们是在一个如岛屿或岛链之类的孤立地域所演化的。在那里,带有花朵的植物可以发展出和某些特定动物(如黄蜂)有着高度特定的关系,而这即是许多岛内物种发展至现今的方式。此一共生关系-假想黄蜂夹带着花粉由一株植物到另一株植物身上,如同今日的无花果小蜂一般,最终会导致植物和其共生同伴间高度的特化。岛屿遗传学被认为是物种形成的主要来源,尤其是当其演进似乎需要有次要的过渡形式时。注意,黄蜂并不是一个特例;蜜蜂明显地和植物演化至特定的共生关系,即源自黄蜂。
同样地,大多数在植物生殖过程中的果实源自于花朵的部份增大。果实通常是利用动物想要吃它的心理,好将其内的种子散播出去。
当许多此类的共生关系仍然无法和陆生动物竞争和散播时,花朵表现出其不同的有效生殖方式,并散播(不论其真实来源)出去成为陆生植物的优势类型。
当只有某些艰难的证据显示花朵已存在了1.3亿年之久时,却有一些环境上的证据显示它们已存在了2.5亿年的时间。一种被植物用来保护其花朵的化学物质-齐墩果烷在如此古老的化石植物上被发现到,如大羽羊齿植物其在彼时演化,且带有现代开花植物的许多特征,虽然它们并不知道是否为开花植物,因为只有其茎棘被保留细部地被发现;其为硅化木最早的一个例子。
叶子和茎在构造上的相似非常重要,因为花朵在基因上只是对植物茎叶的适应变化,一个基因的组合通常会对应至一个新芽的形成。大部份的花朵被认为都曾有着不定数量的花朵部份,每一部份通常都和其他部份相区隔(或有关联)。花朵也都曾顷向于长成螺旋的图样,为双性的(在植物内,这是指同一朵花上同时有着雄性和雌性),且子房(雌性部份)占了绝大部份。当花朵更演进之后,某些种类的部份混合在一起,且会有着特定的部份数量和构造,并在每个花朵或植物上有着特定的性别,或至少为“子房下位(Ovary inferior)”。
花朵持续进化至今日;现今的花朵是如此深刻地影响着人类,以致于某些不能在自然界中授粉。如今许多观赏用的花朵也曾是些常见的杂草,只在地面被扰乱时发芽。某些偏向于和人类的稻谷长在一起,或许之间都已经有着共生的关系,且最漂亮的草因为其美丽而不会被拔起,因而发展出一特殊的人择(artificial selection)形态。
被子植物又名瓶状辽宁果
被子植物又称有花植物。包括人类在内的哺乳动物是随着被子植物的兴起而繁盛,并进化发展到如今的高级阶段,因而被子植物的起源和早期演化长期以来一直是古植物学家研究的焦点。科学家先后在朝阳市北票地区的义县组地层中发现了梁氏朝阳序、辽宁古果、中华古果、始花古果、十字中华果、迪拉丽花等被子植物化石。瓶状辽宁果和梁氏朝阳序、辽宁古果、迪拉丽花一样,都是在辽西 北票地区黄半吉沟发现的。瓶状辽宁果保存的是该植物的果实,但科学家可以推断出它的心皮属于瓶状类型。产出这一化石的地层在地质学上称为义县组,属于早白垩世,形成于1.25亿年前。王鑫称:这么多被子植物化石在早白垩世义县组的出现再次提示人们,早白垩世不会是被子植物起源的时代,否则就会非常难于解释被子植物的爆发式涌现。
穿越1.25亿年的历史,又一种有花植物惊现朝阳。中科院南京地质古生物研究所王鑫研究员和渤海大学韩刚副教授在刚刚出版的《地质学报》英文版上报道称,他们在这一化石植物上发现了迄今世界上最早的被子植物的瓶状心皮和基生胎座,因而把这种植物命名为瓶状辽宁果。
心皮是植物进化的产物,是被子植物特有的器官。传统的被子植物演化理论认为,像豆荚状的对折心皮是最原始的,而且认为对折心皮中的边缘胎座是最原始的胎座形式。过去对于被子植物的系统学研究表明,瓶状心皮和基生胎座都应当出如今原始的早期被子植物中,而这一观点此前一直缺乏相应的化石证据。瓶状心皮和基生胎座在早白垩世的瓶状辽宁果中的出现是支持上述观点的第一件化石证据。
起源时间
当前多数学者认为被子植物起源于白垩纪或晚侏罗纪。根据斯科特(Scott)、马朗(Ba-rghoon)和利奥波德(Leopold)(1960)对以前记述过的化石进行了全面的讨论,发现白垩纪之前未曾保存具确实证据的被子植物化石。此外从孢粉证据来看,同样表如今白垩纪以前的地层中,未能找到被子植物花粉。多伊尔(Doyle 1969)和马勒(Muller 1970)根据早白垩世和晚白垩世地层之间孢粉的研究,支持被子植物最初的分化是发生在早白垩世,大概在侏罗纪时期就为这个类群的发展准备好了条件,这一观点也被奥尔夫(Wolf 1972)从美国弗吉尼亚的怕塔克森特早白垩世岩层中得到的叶化石证据所支持。同时,他们还得出结论:在白垩纪,木兰目的发展先于被子植物的其它类群。我国学者潘广等人近10年在对华北燕辽地区中侏罗纪地层中发现并确证了原始被子植物的存在,也发现了那时的单子叶和双子叶植物---木兰类和柔夷花序类均已发育较好。因此,被子植物的起源应早于白垩纪。这个观点已在1999年第16届国际植物学大会上引起关注。
发源地
 关于被子植物起源的时间,最好的花粉粒和叶化石证据表明,被子植物出现于1.2~1.35亿年前的早白垩纪。在较古老的白垩纪沉积中,被子植物化石记录的数量与蕨类和裸子植物的化石相比还较少,直到距今8000~9000万年的白垩纪末期,被子植物才在地球上的大部分地区占了统治地位。至于被子植物起源的地点,当前普遍认为被子植物的起源和早期的分化很可能在白垩纪的赤道带或靠近赤道带的某些地区,其根据是现存的和化石的木兰类在亚洲东南部和太平洋南部占优势,在低纬度热带地区白垩纪地层中发现有最古老的被子植物三沟花粉。中国植物分类学家吴征镒教授,从中国植物区系研究的角度出发,提出整个被子植物区系早在第三纪以前,便在古代统一的大陆的热带地区发生,并认为中国南部、西南部和中南半岛,在北纬20°~40°间的广大地区,最富于特有的古老科、属,这些第三纪古热带起源的植物区系,即是近代东亚温带、亚热带植物区系的开端,这一地区就是被子植物的发源地。关于被子植物起源的地点问题,依然处于推测阶段,虽然多数学者赞同低纬度起源,但要确切回答被子植物的起源地点还有困难,有待作更深入的研究。关于被子植物的祖先,推测很多,并无定论。其中有藻类、蕨类、松杉目、买麻藤目、本内苏铁目、种子蕨和舌羊齿等,多数学者认为,应到已绝灭的古老的裸子植物中去寻找被子植物的祖先。比较流行的是本内苏铁和种子蕨这两种假说。
关于被子植物起源的时间,最好的花粉粒和叶化石证据表明,被子植物出现于1.2~1.35亿年前的早白垩纪。在较古老的白垩纪沉积中,被子植物化石记录的数量与蕨类和裸子植物的化石相比还较少,直到距今8000~9000万年的白垩纪末期,被子植物才在地球上的大部分地区占了统治地位。至于被子植物起源的地点,当前普遍认为被子植物的起源和早期的分化很可能在白垩纪的赤道带或靠近赤道带的某些地区,其根据是现存的和化石的木兰类在亚洲东南部和太平洋南部占优势,在低纬度热带地区白垩纪地层中发现有最古老的被子植物三沟花粉。中国植物分类学家吴征镒教授,从中国植物区系研究的角度出发,提出整个被子植物区系早在第三纪以前,便在古代统一的大陆的热带地区发生,并认为中国南部、西南部和中南半岛,在北纬20°~40°间的广大地区,最富于特有的古老科、属,这些第三纪古热带起源的植物区系,即是近代东亚温带、亚热带植物区系的开端,这一地区就是被子植物的发源地。关于被子植物起源的地点问题,依然处于推测阶段,虽然多数学者赞同低纬度起源,但要确切回答被子植物的起源地点还有困难,有待作更深入的研究。关于被子植物的祖先,推测很多,并无定论。其中有藻类、蕨类、松杉目、买麻藤目、本内苏铁目、种子蕨和舌羊齿等,多数学者认为,应到已绝灭的古老的裸子植物中去寻找被子植物的祖先。比较流行的是本内苏铁和种子蕨这两种假说。
可能的祖先
 被子植物的属种十分庞杂,形态变化很大,分布极广,粗看起来,确实难用统一的特征将所有的被子植物归成一类。因此,对被子植物的祖先存在不同的假说,有多元论和单元论两种起源说。
被子植物的属种十分庞杂,形态变化很大,分布极广,粗看起来,确实难用统一的特征将所有的被子植物归成一类。因此,对被子植物的祖先存在不同的假说,有多元论和单元论两种起源说。
多元论认为被子植物来自许多不相亲近的群类,彼此是平行发展的。胡先骕、米塞(Meeuse)、恩格勒(Engler)和兰姆(Lam)等人是多元论的代表。我国的分类学家胡先骕1950年发表了一个被子植物多元起源的系统,也是我国学者发表的被子植物的唯一系统。
单元论是当前多数植物学家主张的被子植物起源说。主要依据是被子植物有许多独特和高度特化的性状,如雄蕊都有四个孢子(花粉)囊和特有的药室内层;大孢子叶(心皮)和柱头的存在;雌雄蕊在花轴排列的位置固定不变;双受精现象和三倍体胚乳;以及筛管和伴胞的存在。因此,人们认为被子植物只能起源于一个共同的祖先。哈钦森(Hutchinson)、塔赫他间(Takhtajan)、克朗奎斯特(Cronquist)和 Judd是单元论的主要代表。
被子植物如确系单元起源,那么它究竟发生于哪一类植物呢?推测很多,至今未有定论。人们推测的有:藻类、蕨类、松杉目、买麻藤目、本内苏铁目、种子蕨等。当前比较流行的是本内苏铁目和种子蕨这二种假说。
塔赫他间和克朗奎斯特从研究现代被子植物的原始类型或活化石中,提出被子植物的祖先类群可能是一群古老的裸子植物。并主张木兰目为现代被子植物的原始类型。这一观点已得到多数学者的支持。那么,木兰类是从哪一群原始被子植物起源的呢?莱米斯尔(Lemesle)主张起源于本内苏铁,认为本内苏铁的孢子叶球常两性,稀单性,和木兰、鹅掌楸的花相似;种子无胚乳,仅是两个肉质的子叶和次生木质部的构造亦相似等等,从而提出被子植物起源于本内苏铁。但是,上一年来这种见解逐渐减少。塔赫他间认为,本内苏铁的孢子叶球和木兰的花的相似性是表面的,因为木兰类的雄蕊(小孢子叶)像其他原始被子植物的小孢子叶一样是分离、螺旋状排列的,而本内苏铁的小孢子叶为轮状排列,且在近基部合生,小孢子囊合生成聚合囊;其次,本内苏铁目的大孢子叶退化为一个小轴,顶生一个直生胚珠。因此要想象这种简化的大孢子叶转化为被子植物的心皮是很困难的。另外,本内苏铁以珠孔管来接受小孢子,而被子植物通过柱头进行授粉,所有这些都表明被子植物起源于本内苏铁的可能性较小。塔赫他间认为被子植物同本内苏铁有一个共同的祖先,有可能从一群最原始的种子蕨起源。当前,大部分系统发育学家接受种子蕨作为被子植物的可能祖先,但是由于化石纪录的不完全,这种假说的证实还有待更全面、更深入的研究。
被子植物系统发育重建
人类研究植物分类、进化及系统发育的目的是要揭示植物世界发生、发展、绝灭的历史, 从现存被子植物和化石记录,通过宏观与微观特征的研究重建被子植物的系统发育,建立一个完全自然的系统。
形态特征
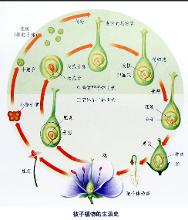
 被子植物的习性、形态和大小差别很大,从极微小的青浮草到巨大的乔木桉树。大多数直立生长,但也有缠绕、匍匐或靠其他植物的机械支持而生长的。多含叶绿素,自己制造养料,但也有腐生和寄生的。有几个科的植物是肉食的,如猪笼草科(Drosera)植物以昆虫和其他小动物为食物。许多是木本的(乔木和灌木),但多为草本,草本被子植物比木本具有更进化的特征。多异花传粉,少数自花传粉。花粉粒到达柱头后即萌发并产生花粉管,通过花柱向下进入子房腔。花粉管前端有管核和生殖细胞。随着花粉管的发育,生殖细胞分裂成两个雄配子。花粉管一般通珠孔进入胚珠。花粉管进入雌配子体后,顶端破裂,两个雄配子释出,进入雌配子体的细胞质内。其中一个雄配子钻入卵细胞内,与之融合而受精。受精后产生的合子常发育成胚。另一个雄配子同雌配子体的其他两个核(极核)结合(或同两极核预先融合成的一个核相结合)形成胚乳核,胚乳核产生胚乳,用以贮藏养料。两个雄配子均参加融合过程,这称为双受精作用,为被子植物所独有。
被子植物的习性、形态和大小差别很大,从极微小的青浮草到巨大的乔木桉树。大多数直立生长,但也有缠绕、匍匐或靠其他植物的机械支持而生长的。多含叶绿素,自己制造养料,但也有腐生和寄生的。有几个科的植物是肉食的,如猪笼草科(Drosera)植物以昆虫和其他小动物为食物。许多是木本的(乔木和灌木),但多为草本,草本被子植物比木本具有更进化的特征。多异花传粉,少数自花传粉。花粉粒到达柱头后即萌发并产生花粉管,通过花柱向下进入子房腔。花粉管前端有管核和生殖细胞。随着花粉管的发育,生殖细胞分裂成两个雄配子。花粉管一般通珠孔进入胚珠。花粉管进入雌配子体后,顶端破裂,两个雄配子释出,进入雌配子体的细胞质内。其中一个雄配子钻入卵细胞内,与之融合而受精。受精后产生的合子常发育成胚。另一个雄配子同雌配子体的其他两个核(极核)结合(或同两极核预先融合成的一个核相结合)形成胚乳核,胚乳核产生胚乳,用以贮藏养料。两个雄配子均参加融合过程,这称为双受精作用,为被子植物所独有。
具有真正的花
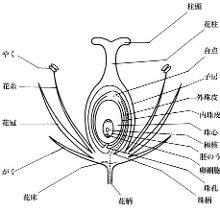
 花是被子植物(被子植物门植物,又称有花植物或开花植物)的繁殖器官,其生物学功能的是结合雄性精细胞与雌性卵细胞以产生种子。这一进程始于传粉,然后是受精,从而形成种子并加以传播。对于高等植物而言,种子便是其下一代,而且是各物种在自然分布的主要手段。同一植物上着生的花的组合称为花序。
花是被子植物(被子植物门植物,又称有花植物或开花植物)的繁殖器官,其生物学功能的是结合雄性精细胞与雌性卵细胞以产生种子。这一进程始于传粉,然后是受精,从而形成种子并加以传播。对于高等植物而言,种子便是其下一代,而且是各物种在自然分布的主要手段。同一植物上着生的花的组合称为花序。
被子植物具异形孢子,亦即能产生两种生殖孢子。 花粉(小孢子,雄性)和胚珠(大孢子,雌性)分别产生于不同器官,但典型的花则同时含有大小孢子,因为它两种器官兼有。
花可视为节间缩短并具繁殖能力的茎的变态,而其节结构也可看作是叶的高度变态。从本质上说,花的结构是由顶端分生组织的花芽和“体轴”分化形成的。花可以以多种方式着生于植物上。如果花没有任何枝干,而是单生于叶腋,即称为无柄花,而其他花上与茎连接并起支持作用的小枝则称为花柄。若花柄具分支且各分支均有花着生,则各分支称为小梗。花柄的顶端膨大部分称为花托,花的各部分轮生于花托之上,四个主要部分从外到内依次是:
 花萼: 位于最外层的一轮萼片,通常为绿色,但也有些植物的呈花瓣状。
花萼: 位于最外层的一轮萼片,通常为绿色,但也有些植物的呈花瓣状。
花冠:位于花萼的内轮,由花瓣组成,较为薄软,常有颜色以吸引昆虫帮助授粉。
雄蕊群:一朵花内雄蕊的总称,花药着生于花丝顶部,是形成花粉的地方,花粉中含有雄配子。
雌蕊群:一朵花内雌蕊的总称,可由一个或多个雌蕊组成。组成雌蕊的繁殖器官称为心皮,包含有子房,而子房室内有胚珠(内含雌配子)。一个雌蕊可能由多个心皮组成,在这种情况下,若每个心皮分离形成离生的单雌蕊,即称为离心皮雌蕊,反之若心皮合生,则称为复雌蕊。雌蕊的黏性顶端称为柱头,是花粉的受体。花柱连接柱头和子房,是花粉粒萌发后花粉管进入子房的通道。

虽然上述结构体现了花的“典型”构造,但自然中各植物的实际构造具有很大的差异性。这些差异对于被子植物的进化具有重大意义,并被植物学家广泛用于确立各植物的种间关系。例如,被子植物的两个亚纲可以通过其花瓣数加以区分:双子叶植物通常有4或5(或者4或5的倍数)片花瓣,而单子叶植物多为3或3的倍数。
大多数植物的花都如上所述,同时具有雌蕊和雄蕊,这在植物学上称为“完全花”、“两性花”或者“雌雄同花”。不过,也有一些植物的花是“不完全花”或“单性花”,即只有雄蕊或雌蕊的花。此种情况下,如果雌花与雄花分别生长在不同的植株上,则称为“雌雄异株”。相反,如果单性的雄花和雌花同生于一植株,则称为“雌雄同株”。
有些植物的花单生于植株上,而有些植物的花则簇生于植株,对于后者而言,这些花若按照一定规律排列于花轴上,便形成了“花序”。在这一点上,必须要注意“花”的实际概念。从植物学角度看,一朵菊花或向日葵并不是一朵花,而是一头状花序,即由许多小花组成的花序,而且其中的所有小花都具有前文提及的结构。有些花具有辐射对称性,亦即如果其花被以任何角度通过中央轴线一分为二,所得的两半都是对称相等的,称为辐射对称花或整齐花,例如月季和桃花。还有些花只能按一个角度切得两个对称面,则称左右对称花或不整齐花,例如金鱼草和大部分兰花。
 具有雌蕊
具有雌蕊
雌蕊由心皮所组成,包括子房、花柱和柱头3部分。胚珠包藏在子房内,得到子房的保护, 避免了昆虫的咬噬和水分的丧失。子房在受精后发育成为果实。果实具有不同的色、香、味,多种开裂方式;果皮上常具有各种钩、刺、翅、毛。果实的所有这些特点,对于保护种子成熟,帮助种子散布起着重要作用,它们的进化意义也是不言而喻的。
双受精现象,即两个精细胞进入胚囊以后, 1个与卵细胞结合形成合子,另1个与2个极核结合,形成3n染色体,发育为胚乳,幼胚以3n染色体的胚乳为营养,使新植物体内矛盾增大,因而具有更强的生活力。所有被子植物都有双受精现象,这也是它们有共同祖先的一个证据。
孢子体高度发达
 被子植物的孢子体,在形态、结构、生活型等方面,比其他各类植物更完善化、多样化,有世界上最高大的乔木,如杏仁桉(Eucalyptus amygdalina Labill.),高达156米;也有微细如沙粒的小草本如无根萍[Wolffia arrhiza (L.) Wimm.],每平方米水面可容纳300万个个体。有重达25千克仅含1颗种子的果实,如王棕(大王椰子)[Roystonea regia (H.B.K.) O. F.Cook];也有轻如尘埃,5万颗种子仅重0.1克的植物如热带雨林中的一些附生兰;有寿命长达6千年的植物,如龙血树(Dracaena draco L.);也有在3周内开花结籽完成生命周期的植物(如一些生长在荒漠的十字花科植物);有水生、砂生、石生和盐碱地生的植物;有自养的植物也有腐生、寄生的植物。在解剖构造上,被子植物的次生木质部有导管,韧皮部有伴胞;而裸子植物中一般均为管胞(只有麻黄和买麻藤类例外),韧皮部无伴胞,输导组织的完善使体内物质运输畅通,适应性得到加强。
被子植物的孢子体,在形态、结构、生活型等方面,比其他各类植物更完善化、多样化,有世界上最高大的乔木,如杏仁桉(Eucalyptus amygdalina Labill.),高达156米;也有微细如沙粒的小草本如无根萍[Wolffia arrhiza (L.) Wimm.],每平方米水面可容纳300万个个体。有重达25千克仅含1颗种子的果实,如王棕(大王椰子)[Roystonea regia (H.B.K.) O. F.Cook];也有轻如尘埃,5万颗种子仅重0.1克的植物如热带雨林中的一些附生兰;有寿命长达6千年的植物,如龙血树(Dracaena draco L.);也有在3周内开花结籽完成生命周期的植物(如一些生长在荒漠的十字花科植物);有水生、砂生、石生和盐碱地生的植物;有自养的植物也有腐生、寄生的植物。在解剖构造上,被子植物的次生木质部有导管,韧皮部有伴胞;而裸子植物中一般均为管胞(只有麻黄和买麻藤类例外),韧皮部无伴胞,输导组织的完善使体内物质运输畅通,适应性得到加强。
配子体进一步退化
被子植物的小孢子(单核花粉粒)发育为雄配子体,  大部分成熟的雄配子体仅具2个细胞(2核花粉粒),其中1个为营养细胞,1个为生殖细胞,少数植物在传粉前生殖细胞就分裂1次,产生2个精子,所以这类植物的雄配子体为3核的花粉粒,如石竹亚纲的植物和油菜、玉米、大麦、小麦等。被子植物的大孢子发育为成熟的雌配子体称为胚囊,通常胚囊只有7个细胞:3个反足细胞、具有2个极核的中央细胞、2个助细胞、1个卵。反足细胞是原叶体营养部分的残余。有的植物(如竹类)反足细胞可多达300余个,有的(如苹果、梨)在胚囊成熟时,反足细胞消失。助细胞和卵合称卵器,是颈卵器的残余。由此可见,被子植物的雌、雄配子体均无独立生活能力,终生寄生在孢子体上,结构上比裸子植物更简化。配子体的简化在生物学上具有进化的意义。
大部分成熟的雄配子体仅具2个细胞(2核花粉粒),其中1个为营养细胞,1个为生殖细胞,少数植物在传粉前生殖细胞就分裂1次,产生2个精子,所以这类植物的雄配子体为3核的花粉粒,如石竹亚纲的植物和油菜、玉米、大麦、小麦等。被子植物的大孢子发育为成熟的雌配子体称为胚囊,通常胚囊只有7个细胞:3个反足细胞、具有2个极核的中央细胞、2个助细胞、1个卵。反足细胞是原叶体营养部分的残余。有的植物(如竹类)反足细胞可多达300余个,有的(如苹果、梨)在胚囊成熟时,反足细胞消失。助细胞和卵合称卵器,是颈卵器的残余。由此可见,被子植物的雌、雄配子体均无独立生活能力,终生寄生在孢子体上,结构上比裸子植物更简化。配子体的简化在生物学上具有进化的意义。
颈卵器消失,其余为卵器
被子植物的上述特征,使它具备了在生存竞争中,优越于其他各类植物的内部条件。被子植物的产生,便地球上第一次出现色彩鲜艳、类型繁多、花果丰茂的景象,随着被子植物花的形态的发展,果实和种子中高能量产物的贮存,使得直接或间接地依赖植物为生的动物界(尤其是昆虫、鸟类和哺乳类),获得了相应的发展,迅速地繁茂起来。
被子植物是植物界进化最高级、种类最多、分布最广、适应性最强的类群。  在不同的系统,被子植物有300多至400多科,1万多属,20-25万种。超过植物界总种数之半。它们分布于各个气候带。由于气温高、雨水多的缘故,热带、亚热带最多。南美亚马逊河区有约4万种。其他热带地区有2-3万种。温带地区因气温降低,雨量少了,种类渐减。以我国情况看,云南省气候条件好,植物达万种以上,而河北省地处北纬26—43度之间。相对种类减少很多,约有2500种。北极地区则大大减少,许多地方几乎无被子植物,仅少数地方有少数种类顽强生存。如北极柳(Salix lanata)、北极罂粟(Papaver radicatum),其分布纬度达80度以上。在南半球南极大陆的莫尔吉特湾詹尼岛附近,有石竹科植物厚叶柯罗石竹(Colobanthus crassifolius)生存。另外,从海拔高度看,地势越高,气温越低,大约每上升100米,气温降低0.5℃,植物种类组成也有变化;在珠穆朗玛峰地区,气候严寒,只有少数耐寒种类方可生存。5000-5500米地区还能找到石竹科的伏繁缕(Stellaria decumbens)。雪莲花(Saussurea involucrata)在新疆天山高处也有分布。
在不同的系统,被子植物有300多至400多科,1万多属,20-25万种。超过植物界总种数之半。它们分布于各个气候带。由于气温高、雨水多的缘故,热带、亚热带最多。南美亚马逊河区有约4万种。其他热带地区有2-3万种。温带地区因气温降低,雨量少了,种类渐减。以我国情况看,云南省气候条件好,植物达万种以上,而河北省地处北纬26—43度之间。相对种类减少很多,约有2500种。北极地区则大大减少,许多地方几乎无被子植物,仅少数地方有少数种类顽强生存。如北极柳(Salix lanata)、北极罂粟(Papaver radicatum),其分布纬度达80度以上。在南半球南极大陆的莫尔吉特湾詹尼岛附近,有石竹科植物厚叶柯罗石竹(Colobanthus crassifolius)生存。另外,从海拔高度看,地势越高,气温越低,大约每上升100米,气温降低0.5℃,植物种类组成也有变化;在珠穆朗玛峰地区,气候严寒,只有少数耐寒种类方可生存。5000-5500米地区还能找到石竹科的伏繁缕(Stellaria decumbens)。雪莲花(Saussurea involucrata)在新疆天山高处也有分布。
衍生
开花植物的花朵是其和其他种子植物间最显著的不同。  花朵让开花植物能够有更广阔的演化关系,且延展了其生态上的利基,以致能使开花植物在陆地上的生态系中称霸。
花朵让开花植物能够有更广阔的演化关系,且延展了其生态上的利基,以致能使开花植物在陆地上的生态系中称霸。
开花植物的雄蕊比起和其相对应的裸子植物的器官要轻小的多,且随着时间使花朵更具多样性,因其对不同特定的授粉方式的适应能力,如某些特殊的授粉媒介。雄蕊亦随着时间演化出了防止自体授粉的机制,更增加了其多样性,让开花植物有着更多的优势。
缩小的雄体
 开花植物雄体的缩小可能是演化来减少其由授粉(即花粉到达母株)至进入子房受精的时间缩短。在裸子植物中,受精的时间可能长达1年之久,而开花植物则可以在授粉后的很短时间内开始其受精的程序。这使得开花植物能够比裸子植物以极快的速度来产生种子。
开花植物雄体的缩小可能是演化来减少其由授粉(即花粉到达母株)至进入子房受精的时间缩短。在裸子植物中,受精的时间可能长达1年之久,而开花植物则可以在授粉后的很短时间内开始其受精的程序。这使得开花植物能够比裸子植物以极快的速度来产生种子。
被子植物紧密的心皮允许其去适应不同的授粉方式,且避免其自体授精的发生,因此维持其多样性的续增。当胚珠受精了之后,心皮和其他周围的组织便会发展成一颗果实,另一个使被子植物可以增加其在陆地生态系优势的机会,以其演化出多样机制的适应能力。心皮和其他补助部份可能会变为果实。
缩小的雌体
缩小的雌体如同缩小的雄体一般,演化成有较快结实的能力,终致开花植物有一年生草本的生命周期,而使其占有更多的利基。
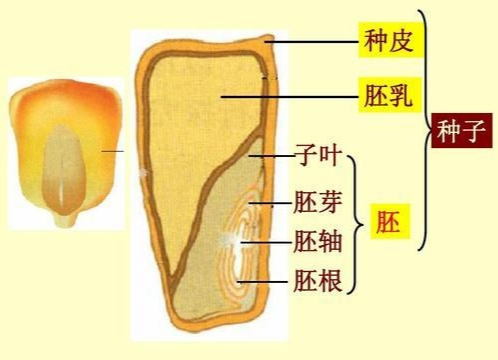 胚乳约在受精后至受精卵第一期前开始成形,有很高的养分可提供胚胎发育之用,且有时亦可提供给幼苗。
胚乳约在受精后至受精卵第一期前开始成形,有很高的养分可提供胚胎发育之用,且有时亦可提供给幼苗。
这些不同的特征一齐使得被子植物为最多样性且大量的陆生植物,并且也对人类最有经济重要性。开花植物在陆地生态系中的优势,有个重大的例外,即在针叶林里。
功能
真正的花为被子植物独具的主要特征,所以被子植物又叫有花植物。花基本上由4个系列的成分组成:
②内层系列为由花瓣组成的花冠,通常质地柔软多汁,色泽鲜艳,具有引诱传粉者的作用;
被子植物群,通过子房上的花柱和柱头接受花粉粒(雄配子体)。花粉萌发后,雄配子体有1个粉管细胞和2个精子(雄核);在胚珠中,雌配子体(胚囊)通常有8个细胞(1个卵细胞、2个助细胞、3个反足细胞、2个极核细胞)。双受精后,由一个花粉粒产生的雄核(配子)与卵受精发育成胚,另一个雄核与二个雌核结合发育成胚乳。花的样式和不同纲的传粉者的感觉或知觉作用紧密相连,而且在昆虫与花的相互关系上存在着平行的协同进化。原始的叶状心皮通过折叠和边缘或缝线的愈合而封闭很可能与虫媒授粉有关,这不仅可以保护胚珠免受攫食昆虫的侵食,而且可以利用来访昆虫作为传递花粉的媒介。
被子植物细胞的结构和分化水平也是最进化的,除了若干原始的成员外,在水分输导组织(木质部)中都有称为导管的管状细胞。在体型上,被子植物大小的变化从高达150米的澳大利亚的桉树到长不足1毫米的、结构简单的微粒状漂浮水生植物无根萍。在热带雨林中,巨大的藤本植物(如榼藤子)攀援而上高耸云霄;也有附生在大树上的兰科、天南星科和凤梨科植物,它们仅依靠树槎上的薄薄积土而生长;茅膏菜、捕蝇草、狸藻和猪笼草等食虫植物则在捕虫设计上结构巧妙和复杂。被子植物中还有各种寄生植物如槲寄生、菟丝子,靠从别的植物中吸取营养物质来生活。特别是分布在加里曼丹和苏门答腊的奇异的大花草,营养器官退化到只剩下几根生长在其寄主植物的根或茎上的丝状物,但它那巨大的花,直径却达45厘米,重7千克,而且充满着为蝇类所喜爱的尸臭味。还有原产于中美洲和南美洲荒漠的多浆汁的仙人掌科植物,约有2000种,形态别致,有高达20米的巨大的仙人柱,有直径达1米的笨重的仙人球,也有延地而生,形似游蛇的仙人鞭。干燥的环境使得这些植物特别耐旱:植物体95%以上都是水,茎的外皮坚硬而不通气,叶变成了刺,而且有些种遍体密布毛茸——这一切都是为了防止水分蒸发。
高水平的生理效率和范围广泛的营养体可塑性以及花的多样性使被子植物得以占领几乎所有极端的生境,并使这些生境特征化——森林、草原、沙漠和许多水生生境。被子植物在主要的植物地理区域内形成了一个常由占优势的科、属和种为特征的一系列广泛的生态群落。
繁殖栽培
有性
异体受精植物往往具有许多防止自体受精的机制,即不亲和性系统。根据有无形态效应可把不亲和性系统分为:同形不亲和性系统和异形不亲和性系统。异形不亲和性系统大约涉及24个被子植物的科,花柱异长,特别是两型花柱,是一种主要的异形不亲和性系统,其中种的群体是由具长花柱和一组短雄蕊的花和具短花柱和一组长雄蕊的花的植物所组成,具体例子有报春花属、耳草属、睡菜属、连翘属等。受精作用仅仅发生在两种类型植物之间的传粉以后,而不是在同一植株上。三型花柱(Tristyly)是花柱异长的另一种型式,包括3种不同类型的花:①长花柱型,柱头下方有2组花药;②中等花柱型,柱头的上、下方各有1组花药;③短花柱型,两组花药都在柱头上方。三型花柱的种仅在酢浆草科、雨久花科和千屈菜科内发现。种内传粉后受精的程度以花粉与柱头在同一水平者为最高。因此,如中等花柱型的花只有在由长花柱型或短花柱型花中的中等高度的花药传粉以后才能产生种子。几乎任何一个两型花柱或三型花柱的属内都有一些植物,它们的花药和柱头位于同一水平上,即使不是真正自花传粉的话也是自交可亲和的。这种花柱同长的植物的分布往往比其花柱异长的植物宽广,如果以自花受精为主,则它们的花可能远较小。
另一类促进异花受精的机制是雌雄异株、雌雄同株,以及它们的各种取代。雌雄异株的分类群可能是从有花植物不同类群的雌雄同株情况下独立发展的——不是直接来自雌雄同株,就是间接通过雌性两性异株,雄性两性异株或雌雄同株等中间阶段而来。通常认为雌雄异株或可以导致雌雄异株的各种中间阶段的进化是有利于异型杂交的选择压力引起的,但还可能包括许多其他的因素。过去曾错误地认为绝大多数雌雄异株的被子植物是风媒传粉的,现 在知道它们大多数是以动物为传粉媒介的。在分类学上,要把雌雄异株种的雄株和雌株配在一起有时很困难。相反,在酸模属、玉叶金花属和羊蹄甲属等植物中,性的这种分离只有为分类提供用来区分近缘种的有用的鉴别依据。
自体受精植物又叫近亲繁殖植物。由于这类植物的种内或分类群内个体的基因型都多少不相同,而每一个个体又能保持其遗传性多代不变,结果往往形成许多纯系或同形小种。闭花受精是一种有利于自体受精的现象,植物的形小而不引人注目的花在花期保持不开放并进行自花传粉和受精。在通常情况下,闭花受精花与开花受精的花生在同一植株上,例如堇菜属、酢浆草属、胡枝子属、活血丹属、野芝麻属、四棱草属和莸属。闭花受精有时与生态条件紧密相关,长时期的多雨天气如同极度荫蔽一样,似有利于产生闭花受精的花(如宝盖草)。这可能是因为在遮蔽状况下传粉昆虫常变得稀少的缘故。堇菜属的闭花受精在高海拔地区较频繁的事实显然应归于光周期反应。福斯卡尔鸭跖草生长在地下的闭花受精的花在曝光条件下可变成开花受精。J·S·赫克斯利认为闭花受精的形成是由生态压力引起的花不能开放(假闭花受精)结果导致自花传粉,继之以花瓣和雄蕊体积的缩小和最后花粉在原来位置上萌发而不从花药上释放等的进一步适应。
二型花严格说来是指闭花受精的花与开花受精的花分别生长在不同的植株上。但闭花受精和二型花现象有时不易分开。例如,在水金凤中,除了完全着生小形的闭花受精花的植株和完全着生形大美丽的开花受精花的植株外,有些植株两者兼备。有时二型花植株在群体内呈多态现象,如卡罗来纳紫草。
无性
无融合生殖植物包括任何类型的无性繁殖。可分为两大类:营养繁殖和无融合结籽。
营养繁殖又叫营养体无融合生殖,是指完全靠匍匐茎、根状茎、块茎、珠芽和冬芽等营养体传代的生殖方式,是由植物体的根、茎、叶等营养器官或某种特殊组织产生新植株的生殖方式。这种生殖方式不涉及性细胞的融合,所以是无融合生殖的一种方式,属于广义的无性生殖范畴。如果人为地取下植物体的部分营养器官或组织,在离体条件下培养成新植株,则称人工营养繁殖。
植物有性繁殖的后代具备双亲的遗传特性容易发生变异,而营养繁殖则不然,如高度杂合的木本多年生植物(如果树),通过人工营养繁殖可保持母本的优良遗传性状。营养繁殖实质上是通过母体细胞有丝分裂产生子代新个体,后代一般不发生遗传重组,在遗传组成上和亲本是一致的。
不同植物类群,有不同的繁殖方式。低等植物通过孢子(无性孢子)或植物体碎片和裂片形成新个体。有些苔藓植物表面可以产生一种特殊器官——胞芽杯,由它长出绿色胞芽。胞芽成熟后从植物体上脱落,遇到适当条件便可长成新的配子体。
种子植物的茎段,是多数植物繁殖的有效器官。例如,草莓属的匍匐茎,即一种细长而沿着地表生成的茎,是从莲座状叶腋中长出,它的每个节都可以长出新的植株。用这种方式繁殖的还有蛇莓、狗芽根、白三叶草、筋骨草和虎耳草等。许多草本多年生植物可通过变态的茎繁殖,如鳞茎、球茎、块茎和根状茎,这些变态茎具有贮藏食物的功能,也是营养繁殖的器官。鳞茎实际上是短而膨大的竖立苗端,肉质叶鳞包围其生长点和花原基。由叶鳞腋间产生小鳞茎,最终脱离母鳞茎形成新植株。这种繁殖方式见于洋葱、水仙、郁金香、风信子、百合、大蒜和贝母等。有的百合叶腋可以长出零余子,即小鳞茎,又叫珠芽,它脱离母体后可以长成一个新植株。唐菖蒲、藏红花和小苍兰贝的茎是球茎。唐菖蒲球茎上有4个芽原基,这些芽原基在适当条件下可以发育形成新球茎,以后老球茎开花后死亡。在每个新球茎周围,又可长出一些大小不同的小球茎,当他们生长1~2年以后也可达到开花阶段。块茎为肉质地下茎缩短膨大的产物,把具芽眼的马铃薯块茎切成小块栽培时,从芽眼可长出苗,再由苗端下部长出不定根。根状茎是地下水平生长的主茎,具节和节间,叶、花轴和不定根等可从节上发生。如鸢尾、美人蕉、竹子和有毒杂草、阿拉伯高粱等都有根状茎。许多重要经济植物如香蕉、姜、蕨类和某些禾本科植物也是靠根状茎繁殖的。
根是营养繁殖的另一种重要器官。例如:玫瑰、杨树、覆盆子和悬钩子等植物的水平根系上可产生不定芽(根出条),并可以陆续发育出新植株。每个带新根的苗都可移植成株。块根为膨大肉质的变态地下根,如果把白薯可食部分的块根放在苗床上,可长出不定芽,由不定芽茎部长出不定根,种植后由不定根又可膨大形成新的块根。
叶和芽也能繁殖。如落地生根的肉质叶缘每一缺刻都能产生“胚”,这种胚发育到一定程度,小苗就可落地生根,并发育为独立的新植株。过山蕨和鞭叶铁线蕨的叶轴顶端尖细,并延伸成鞭状,着地后即可生根,长出新的植株。浮萍和凤眼蓝等水生植物还可由叶子茎部的侧芽产生新植株。龙舌兰在开花死后,新植株可由老叶的腋芽产生。
著名的例子有伊乐藻(只存在一个性别的雌雄异株的种)、水剑叶(Stratiotes aloides)、黑藻等以及浮萍科的某些种,它们在北欧完全是营养繁殖,在别处却是正常的有性繁殖。假胎生现象是一种繁殖体发生在花内部而且代替了花的营养繁殖方式,在虎耳草属、龙舌兰属、葱属、蓼属以及禾本科的早熟禾属和羊茅属等属内很有名。但其中有些种在同一花序上兼有有性的和假胎生的花(如拳参和薤白)。
⑵无融合结籽
无融合结籽包括用无性方法产生胚胎和种子的任何类型,其特点是绕过减数分裂和受精,因此最后形成的胚胎的染色体数目和基因型与母株完全一样。大致有以下几种方式:①不定胚生殖。胚胎直接由作为二倍性孢子体母体组织的珠心或珠被产生,完全避开配子体阶段。以柑橘属为最著称;还发生在冬青叶山麻杆、甜味大戟、齿叶金莲木、蒲桃、桃叶野扇花、橙黄仙人掌,以及玉簪属、葱属和绶草属等植物内;②无孢子生殖和双倍孢子生殖。前者是由珠心或内珠被营养细胞经过多次体细胞分裂而直接产生胚囊;后者是胚囊虽由大孢子母细胞产生,但产生过程中或则根本没有减数分裂,或则减数分裂大为变样,以致染色体不进行配对或减数。从形态学观点看,这两种无融合生殖方式仍有孢子体与配子体的世代交替,但因绕过减数分裂而使配子也是二倍性的;③假受精。通过授粉作用与花粉管发生使卵受雄配子的刺激后形成种子,但雄核绝不与卵融合,所以种子后代的基因型与母株相同。
无融合生殖按其在个体发生上的不同程度又可分为专性无融合和兼性无融合两大类。前者是指整个植株完全是无融合生殖的(如大蒜);后者是指同一植株上既有无融合生殖又有有性生殖(如薤白和拳参)。由于兼性无融合的有性过程能产生一系列新的无性系,从而在新、老无性系之间形成一种异常复杂的关系:虽有稳定的性状区别,但这种区别非常微小,以致很难作为分种的依据。再有,因无融合生殖常与种间杂交和多倍化现象密切相关,使变异式样更为复杂,形成被称为“无融合复合种”的,分类学上十分困难的类群,如还阳参属、早熟禾属、委陵菜属、悬钩子属、银胶菊属、蒲公英属和山柳菊属等。
种类介绍
被子植物分为2个纲,即双子叶植物纲和单子叶植物纲,它们的基本区别如下:
| 分类 | 双子叶植物纲 | 单子叶植物纲 |
|---|---|---|
| 子叶 | 具2片子叶 | 具1片子叶 |
| 根 | ,直根系 | ,须根系 |
| 茎 | 维管束成环状排列,有,形成层 | 维管束成星散排列,无形成层 |
| 叶 | 具,网状脉 | 具平形脉或弧形脉 |
| 花 | 各部分基数为4或5花粉粒具3个萌发孔 | 各部分基数为3, 花粉粒具单个萌发孔 |
| 胚 | 具2片子叶 | 具1片子叶 |
以上区别点不是绝对的,实际上有交错现象,如双子叶植物纲中的毛茛科、车前科、菊科等有须根系植物;胡椒科、睡莲科、毛茛科、石竹科等有维管束星散排列的植物;樟科、木兰科、小檗科、毛茛科有3基数的花;睡莲科、毛茛科、小续科、罂粟科、伞形科等有 l片子叶的现象。单子叶植物纲中的天南星科、百合科、薯蓣科等有网状脉;眼子菜科、百合科、百部科等有4基数的花
恩格勒系统
这是德国分类学家恩格勒(A.Engler)和勃兰特(K.PrantI)于1897年在其《植物自然分科志》巨著中所使用的系统,  它是分类学史上第一个比较完整的系统,它将植物界分13门,第13门为种子植物门,再分为裸子植物和被子植物两个亚门,被子植物亚门包括单子叶植物和双子叶植物两个纲,并将双子叶植物纲分为离瓣花亚纲(古生花被亚纲)和合瓣花亚纲(后生花被亚纲)。
它是分类学史上第一个比较完整的系统,它将植物界分13门,第13门为种子植物门,再分为裸子植物和被子植物两个亚门,被子植物亚门包括单子叶植物和双子叶植物两个纲,并将双子叶植物纲分为离瓣花亚纲(古生花被亚纲)和合瓣花亚纲(后生花被亚纲)。
恩格勒系统将单子叶植物放在双子叶植物之前,将合瓣花植物归并一类,认为是进化的一群植物,将柔荑花序植物作为双子叶植物中最原始的类群,而把木兰目、毛茛目等认为是较为进化的类群,这些观点为现代许多分类学家所不赞同。
思格勒系统几经修订,在1964年出版的《植物分科志要》第十二版中,已把双子叶植物放在单子叶植物之前。共有62目,344科,其中双子叶植物48目,290科,单子叶植物14目,54科。药用植物学教材被子植物分类部分采用修订的思格勒系统,但有的内容有变动。
哈钦松系统
英国植物学家哈钦松(J.Hutchinson)于1926年和1934年在其《有花植物科志》I、 II中所建立的系统。  在1973年修订的第三版中,共有111目,411科,其中双子叶植物82目,342科,单子叶植物29 目,69科。
在1973年修订的第三版中,共有111目,411科,其中双子叶植物82目,342科,单子叶植物29 目,69科。
哈钦松系统认为多心皮的木兰目、毛茛目是被子植物的原始类群,但过分强调了木本和草本两个来源,认为木本植物均由木兰目演化而来,草本植物均由毛茛目演化而来,结果使得亲缘关系很近的一些科在系统位置上都相隔很远,如草本的伞形科和木本的山茱萸科、五加科;草本的唇形科和木本的马鞭草科等,这种观点亦受到现代多数分类学家所反对。
塔赫他间系统
前苏联植物学家 塔赫他间(A.Takhtajan)  于 1954年在其 《被子植物起源》一书中公布的系统,他首先打破了传统把双子叶植物分为离瓣花亚纲和合瓣花亚纲的分类;在分类等级上增设了“超目”一级分类单元。他将原属毛茛科的芍药属独立成芍药科等,都和当今植物解剖学、孢粉学、植物细胞 分类学和化学分类学的发展相吻合,在国际上得到共识。
于 1954年在其 《被子植物起源》一书中公布的系统,他首先打破了传统把双子叶植物分为离瓣花亚纲和合瓣花亚纲的分类;在分类等级上增设了“超目”一级分类单元。他将原属毛茛科的芍药属独立成芍药科等,都和当今植物解剖学、孢粉学、植物细胞 分类学和化学分类学的发展相吻合,在国际上得到共识。
塔赫他间系统经过多次修订,在1980年修订版中,共有28超目,92目,416科,其中双予叶植物(木兰纲)20超目,71目,333科,单子叶植物(百合纲)8超目,21目,77科,显得较繁锁。
克朗奎斯特系统
美国学者阿瑟·克朗奎斯特(1919年-1992年)最早于1958年发表的一种对有花植物进行分类的体系,  1981年在他的著作《有花植物的综合分类系统》中最终完善。包括64个目和383个科,是数个常被使用的被子植物分类系统之一。
1981年在他的著作《有花植物的综合分类系统》中最终完善。包括64个目和383个科,是数个常被使用的被子植物分类系统之一。
克朗奎斯特系接近于塔赫他间系统,把被子植物门(称木兰植物门)分成木兰纲和百合纲,但取消了“超目”一级分类单元,科的划分也少于塔赫他间系统。
现中国有的教科书采用这一系统。
被子植物种系发生学组
") 被子植物种系发生学组(APG,Angiosperm Phylogeny Group)是一个非正式的国际植物分类学组织,这个组织试图将分子生物学原理应用到显花植物的分类中,以求得能为大多数学者所共识的分类方法。被子植物种系发生学组至今发表三篇论文,陈述被子植物的分类系统:
被子植物种系发生学组(APG,Angiosperm Phylogeny Group)是一个非正式的国际植物分类学组织,这个组织试图将分子生物学原理应用到显花植物的分类中,以求得能为大多数学者所共识的分类方法。被子植物种系发生学组至今发表三篇论文,陈述被子植物的分类系统:
《被子植物APG分类法》是1998年由被子植物种系发生学组出版的一种对于被子植物的现代分类法。这种分类法和传统的依照形态分类不同,是主要依照植物的三个基因组DNA的顺序,以亲缘分支的方法分类,包括两个叶绿体和一个核糖体的基因编码。虽然主要依据分子生物学的数据,但是也参照其他方面的理论,例如将真双子叶植物分支和其他原来分到双子叶植物纲中的种类区分,也是根据花粉形态学的理论。
分类系统
经典分类
被子植物在阶层系统中的地位和名称意见不一,有的作为门,有的作为亚门或纲。除了少数分类系统外,被子植物通常被分成双子叶植物和单子叶植物两大部分,但不同的分类系统给予这两大类群以不同的等级和名称——有的作为纲,即双子叶植物纲和单子叶植物纲,有的则作为亚纲,即双子叶植物亚纲(Dicoty-ledonidae)和单子叶植物亚纲(Monocotyledonidae)。这两大类群彼此以若干性状相区别,但孢粉学资料并未提供它们之间存在着明显界线的证据。分支系统学家则认为单子叶植物和双子叶植物是非单元发生的并系类群,在自然分类系统中应予排除。
亚纲又进一步被分为超目和目。超目这个介于亚纲和目之间的分类阶元已被现代的被子植物分类系统普遍采用。
科是植物学家和非行家最常用的高级分类阶元。但被子植物科的数目依不同的分类系统及各学者对科的范围看法不同而有较大的变化,一般在300~450之间,1987年A·л·塔赫塔江画分为533个科。被子植物科的内容变化也很大,从只含一个属和一个种的单型科如昆栏树科(Trochodendraceae)和杜仲科(Eucommiaceae)到属以百计和种以千计的庞大的多型科如菊科(Asteraceae)、大戟科(Euphorbiaceae)和茜草科(Rubiaceae)。尽管科的数目和内容有较大的变化,但学者基本一致的看法是大约有200左右的科是“核心的科”。
P·H·戴维斯和J·克伦的《有花植物科的鉴定》第二版(1979年)、R·基辛格等修订的《桑纳有花植物科解析检索表》(1981年)以及A·克朗奎斯特的《有花植物的综合分类系统》(1981年)载有迄今最完备的世界有花植物的分科检索表。然而,目(甚至超目)在不同的分类系统不仅数目相差悬殊,而且名称和内容大相径庭。目除了用在分类系统的正规描述中外,并非广泛使用。许多分类学家喜欢非正式地应用科以上水平的分类学单位。
现代分类
现代的被子植物分类系统并不十分关心什么类群应予承认和应放在什么等级上,所关心的是不同科之间的亲缘关系以及如何在一个系统方案或排列中,反映出科的系统发育或进化的状况和起源。但20世纪60年代以来,学者在以下两个问题上存在着很大的分歧:
②评价分类群之间的亲缘关系是根据性状的相似性程度还是通过综合分析性状的进化趋势。分支系统学和进化分类学的学者主张系统发育是生物分类学研究的目标和建立分类系统的依据,和用系统树来表示系统发育的关系。
所谓系统发育的分类系统是指分类群按阶层系统体系的排列,既为了表示多层次分枝的顺序即进化枝,又为了表示分枝的进化性变化的程度和性质以及它们的级度即进化级。鉴于现有被子植物的化石很不充分,因此,反映被子植物系统发育关系的系统图,只能来自对现存类型比较研究的演绎。但以J·S·L·吉尔默与V·H·海吾德为代表的表征分类学学者强调,鉴于企图通过完整的化石证据来重建被子植物系统发育历史的愿望不可能有任何把握实现,因此“被子植物的分类应该表达植物之间的关系,像它们存在的那样,而且应该以全面的相似性来评价”,而“推测性的系统发育不应该作为分类的基础”。具体地说,他们主张用最大数量的属性,以全面的相似性和区别性为依据进行通用的分类,以便有可能为更多的目的服务(边缘学科,使用数学方法和电子计算机研究解决生物学中的分类问题。也称数值分类学)。
分类
自达尔文始,已有30多位学者提出了被子植物的(有的是种子植物或维管植物的)分类系统。但由于被子植物的系统发育中普遍存在的:
①镶嵌进化或异级度性;
②趋同进化;
③生殖阻障的起源和表征的分化之间存在着某种独立性;
④由杂交、异源多倍化、遗传物质的非谱系传递(如通过病毒)、以及很不相同的分类群的共生性“融合”所造成的非趋异性网状进化,使这项工作的复杂性大大地增加了。
但一般对被子植物的单元发生、单子叶和双子叶植物间的亲缘关系、木兰目和与其近缘目的原始性、具退化单性花的风媒传粉科的次生性质等许多问题,现已基本上取得了一致的认识。
几个现代著名的被子植物分类系统有许多不一致的地方,这不仅有形式方面的问题,也有基本原理方面的问题。例如,在有关分类群的范围方面,有的使用范围较大的高级分类群来更好地表示亲缘关系(如R·F·索恩系统,1983年;A·克朗奎斯特系统,1981年),有的则使用范围较小、特征较明显的、无疑具有共同祖先的高级分类群(如R·达尔格伦系统,1983年;A·л·塔赫塔江系统,1980年、1988年)。克朗奎斯特系统着重于形态性状和实际应用方面,检索表一直到科。对比之下,达尔格伦系统更重视化学、超微结构和胚胎学性状,显然不那么容易被野外和标本室分类学家所理解。
A·克朗奎斯特(1981年)分类系统的系统大纲是:将被子植物门(Angiospermae,又称木兰门,Magnoliophyta)分为双子叶植物纲(Dicotyledoneae,又称木兰纲,Magnoliopsida)和单子叶植物纲(Monocotyledoneae,又称百合纲Liliopsida)。双子叶植物纲分为6个亚纲:木兰亚纲(Magnoliidae)、金缕梅亚纲(Hamamelidae)、石竹亚纲(Caryophyllidae)、五桠果亚纲(Dilleniidae)、蔷薇亚纲(Rosidae)和菊亚纲(Asteridae);将单子叶植物纲分为5个亚纲:泽泻亚纲(Arismatidae)、鸭跖草亚纲(Commelinidae)、槟榔亚纲(Arecidae)、姜亚纲(Zingiberidae)和百合亚纲(Liliidae)。
A·克朗奎斯特的基本观点是:
②现代所有生活的被子植物各亚纲都不可能是从现存的其他亚纲的植物进化来的;
③木兰亚纲是被子植物基础的复合群,也就是通常所称的毛茛复合群,花被十分发育,雄蕊多数、向心发育,具2核花粉和单淘花粉,雌蕊由单心皮组成,2层珠被,厚珠心胚珠,除樟科外都具内胚乳,它是被子植物中原始的一个亚纲,木兰目(Magnoliales)是现存的最原始的被子植物;
④金缕梅亚纲是一群花减化(无瓣、生在柔荑花序上)的风媒传粉群,在将一些无关的科如杨柳科(Salicaceae)移出之后,这个亚纲主要还是传统的“柔荑花序类”植物;
⑤石竹亚纲是由石竹目和与该目有直接亲缘关系的类群组成,这个群趋向于具有特立中央胎座或基底胎座,许多成员都含有甜菜碱,这在其他亚纲中是不存在的,该亚纲仅有几个科是合瓣的;
⑥蔷薇亚纲的成员具有多数雄蕊,雄蕊向心发育,比较进步的成员强烈地趋向于具单胚珠的室和由于一轮雄蕊退化而形成蜜腺盘,绝大多数是离瓣的,只有少数成员是合瓣的或无瓣的;
⑦五桠果亚纲具有多数雄蕊,雄蕊离心发育,通常每室的胚珠多于一枚,偶尔有由雄蕊起源的蜜腺盘,该亚纲较进化的科中有合瓣的成员;
⑧菊亚纲包括了进化较高级的合瓣科,雄蕊数很少多于花冠的裂片数,具薄珠心胚珠和一层珠被;
⑨单子叶植物起源于类似现代睡莲目(Nymphaeales)的双子叶植物,这群植物为水生、花具有离生心皮、具有普通的花被、单孔花粉、片状胎座、失去或一开始就不具形成层和导管系统等,因此认为泽泻亚纲是单子叶植物基础的一个类群;
⑩单子叶植物的5个亚纲开拓了不同的生态龛或一组生态龛:泽泻亚纲主要是水生的,其他亚纲则主要是陆生的;典型的槟榔亚纲植物有大而通常具柄的叶,常为乔木状、花聚生成肉穗花序,除天南星目外,有很发达的导管系统,棕榈类是该亚纲的顶峰;鸭跖草亚纲开拓了花的退化和风媒传粉的途径,一直发展到禾本科和莎草科;姜亚纲的绝大多数成员分布于热带,既有陆生也有附生,以花具蜜腺和上位花(子房下位)、花整齐或不整齐而不同于其他单子叶植物;百合亚纲开拓了高度发展的虫媒传粉途径,具花瓣状的萼片和花瓣、合生心皮,大多数成员是陆生或附生草本,常有鳞茎、块茎或球茎,而兰科是发展的顶峰。
克朗奎斯特系统(1981修订版)完整目录如下
1.Magnoliopsida(木兰纲) | 1.木兰亚纲(Magnoliidae) | 1.木兰目(Magnoliales) | 1.林仙科(Winteraceae) |
2.单室木兰科(Degeneriaceae) | |||
4.帽花木科(Eupomatiaceae) | |||
5.木兰藤科(Austrobaileyaceae) | |||
6.木兰科(Magnoliaceae) | |||
7.短蕊花科(Lactoridaceae) | |||
9.肉豆蔻科(Myristicaceae) | |||
10.白樟科(Canellaceae) | |||
1.无油樟科(Amborellaceae) | |||
2.早落瓣科(Trimeniaceae) | |||
3.杯轴花科(Monimiaceae) | |||
4.腺蕊花科(Gomortegaceae) | |||
5.腊梅科(Calycanthaceae) | |||
6.奇子树科(Idiospermaceae) | |||
7.樟科(Lauraceae) | |||
8.莲叶桐科(Hernandiaceae) | |||
1.金粟兰科(Chloranthaceae) | |||
2.三白草科(Saururaceae) | |||
3.胡椒科(Piperaceae) | |||
1.马兜铃科(Aristolochiaceae) | |||
1.八角科(Illiciaceae) | |||
2.五味子科(Schisandraceae) | |||
1.莲科(Nelumbonaceae) | |||
2.睡莲科(Nymphaeaceae) | |||
3.合瓣莲科(Barclayaceae) | |||
4.莼菜科(Cabombaceae) | |||
5.金鱼藻科(Ceratophyllaceae) | |||
1.毛茛科(Ranunculaceae) | |||
2.星叶草科(Circaeasteraceae) | |||
3.小檗科(Berberidaceae) | |||
5.木通科(Lardizabalaceae) | |||
6.防己科(Menispermaceae) | |||
7.马桑科(Coriariaceae) | |||
8.清风藤科(Sabiaceae) | |||
1.罂粟科(Papaveraceae) | |||
2.紫堇科(Fumariaceae) | |||
1.昆栏树目(Trochodendrales) | 1.水青树科(Tetracentraceae) | ||
2.昆栏树科(Trochodendraceae) | |||
1.连香树科(Cercidiphyllaceae) | |||
2.领春科(Eupteleaceae) | |||
3.悬铃木科(Platanaceae) | |||
5.香灌木科(Myrothamnaceae) | |||
1.虎皮楠科(Daphniphyllaceae) | |||
4.对药树目(Didymelales) | 1.双颊果科(Didymelaceae) | ||
1.杜仲科(Eucommiaceae) | |||
1.钩毛树科(Barbeyaceae) | |||
2.榆科(Ulmaceae) | |||
3.大麻科(Cannabaceae) | |||
4.桑科(Moraceae) | |||
5.蚁栖树科(Cecropiaceae) | |||
6.荨麻科(Urticaceae) | |||
7.(Leitneriales) | 1.塞子木科(Leitneriaceae) | ||
1.马尾树科(Rhoipteleaceae) | |||
2.胡桃科(Juglandaceae) | |||
1.杨梅科(Myricaceae) | |||
10.山毛榉目(Fagales) | 1.橡子木科(Balanopaceae) | ||
2.太果木科(Ticodendraceae) | |||
3.壳斗科(Fagaceae) | |||
4.南青冈科(Nothofagaceae) | |||
5.桦木科(Betulaceae) | |||
3.石竹亚纲(Caryophyllidae) | 1.石竹目(Caryophyllales) | 1.商陆科(Phytolaccaceae) | |
2.透镜籽科(Achatocarpaceae) | |||
3.紫茉莉科(Nyctaginaceae) | |||
4.番杏科(Aizoaceae) | |||
5.龙树科(Didiereaceae) | |||
6.仙人掌科(Cactaceae) | |||
7.藜科(Chenopodiaceae) | |||
8.苋科(Amaranthaceae) | |||
10.落葵科(Basellaceae) | |||
11.粟米草科(Molluginaceae) | |||
12.石竹科(Caryophyllaceae) | |||
2.蓼目(Polygonales) | 1.蓼科(Polygonaceae) | ||
3.蓝雪目(Plumbaginales) | |||
4.五桠果亚纲(Dilleniidae) | 1.五桠果科(Dilleniaceae) | ||
2.芍药科(Paeoniaceae) | |||
1.金莲木科(Ochnaceae) | |||
2.球萼树科(Sphaerosepalaceae) | |||
3.旋花树科(Sarcolaenaceae) | |||
4.龙脑香科(Dipterocarpaceae) | |||
5.油桃木科(Caryocaraceae) | |||
6.山茶科(Theaceae) | |||
8.木果树科(Scytopetalaceae) | |||
9.五列木科(Pentaphylacaceae) | |||
10.四籽树科(Tetrameristaceae) | |||
11.假红树科(Pellicieraceae) | |||
12.五蕊茶科(Oncothecaceae) | |||
13.马克木科(Marcgraviaceae) | |||
14.羽叶树科(Quiinaceae) | |||
15.沟繁缕科(Elatinaceae) | |||
16.八蕊树科(Paracryphiaceae) | |||
17.水母柱科(Medusagynaceae) | |||
18.藤黄科(Clusiaceae) | |||
1.杜英科(Elaeocarpaceae) | |||
2.椴树科(Tiliaceae) | |||
3.梧桐科(Sterculiaceae) | |||
4.木棉科(Bombacaceae) | |||
5.锦葵科(Malvaceae) | |||
1.玉蕊科(Lecythidaceae) | |||
1.瓶子草科(Sarraceniaceae) | |||
2.猪笼草科(Nepenthaceae) | |||
3.茅膏菜科(Droseraceae) | |||
6.堇菜目(Violales) | |||
2.围盘树科(Peridiscaceae) | |||
3.红木科(Bixaceae) | |||
4.半日花科(Cistaceae) | |||
5.蒜树科(Huaceae) | |||
6.裂药花科(Lacistemataceae) | |||
7.杯盖花科(Scyphostegiaceae) | |||
9.堇菜科(Violaceae) | |||
10.柽柳科(Tamaricaceae) | |||
11.瓣鳞花科(Frankeniaceae) | |||
12.双钩叶科(Dioncophyllaceae) | |||
13.钩枝藤科(Ancistrocladaceae) | |||
14.时钟花科(Turneraceae) | |||
15.王冠草科(Malesherbiaceae) | |||
16.西番莲科(Passifloraceae) | |||
17.钟花科(Achariaceae) | |||
19.福桂花科(Fouquieriaceae) | |||
20.单柱花科(Hoplestigmataceae) | |||
21.葫芦科(Cucurbitaceae) | |||
22.四数木科(Datiscaceae) | |||
23.秋海棠科(Begoniaceae) | |||
24.刺莲花科(Loasaceae) | |||
7.杨柳目(Salicales) | 1.杨柳科(Salicaceae) | ||
1.烈味三叶草科(Tovariaceae) | |||
4.辣木科(Moringaceae) | |||
5.木犀草科(Resedaceae) | |||
9.肉穗果目(Batales) | 1.环蕊科(Gyrostemonaceae) | ||
2.藜木科(Bataceae) | |||
1.翅萼树科(Cyrillaceae) | |||
2.山柳科(Clethraceae) | |||
3.假石南科(Grubbiaceae) | |||
4.岩高兰科(Empetraceae) | |||
7.鹿蹄草科(Pyrolaceae) | |||
8.水晶兰科(Monotropaceae) | |||
1.山榄科(Sapotaceae) | |||
2.柿树科(Ebenaceae) | |||
3.安息香科(Styracaceae) | |||
4.尖药科(Lissocarpaceae) | |||
5.山矾科(Symplocaceae) | |||
1.假轮叶科(Theophrastaceae) | |||
2.紫金牛科(Myrsinaceae) | |||
5.蔷薇亚纲(Rosidae) | 1.瓣裂果科(Brunelliaceae) | ||
2.牛栓藤科(Connaraceae) | |||
3.船形果科(Eucryphiaceae) | |||
4.火把树科(Cunoniaceae) | |||
5.澳楸科(Davidsoniaceae) | |||
6.毛枝树科(Dialypetalanthaceae) | |||
8.腺毛草科(Byblidaceae) | |||
10.弯药树科(Columelliaceae) | |||
11.茶藨子科(Grossulariaceae) | |||
12.鞘叶树科(Greyiaceae) | |||
13.鳞叶树科(Bruniaceae) | |||
14.四柱木科(Anisophylleaceae) | |||
15.假海桐科(Alseuosmiaceae) | |||
16.景天科(Crassulaceae) | |||
17.土瓶草科(Cephalotaceae) | |||
19.蔷薇科(Rosaceae) | |||
20.沙莓科(Neuradaceae) | |||
21.假黄杨科(Crossosomataceae) | |||
22.金壳果科(Chrysobalanaceae) | |||
23.海人树科(Surianaceae) | |||
24.棒木科(Rhabdodendraceae) | |||
2.豆目(Fabales) | 1.含羞草科(Mimosaceae) | ||
2.苏木科(Caesalpiniaceae) | |||
1.胡颓子科(Elaeagnaceae) | |||
2.山龙眼科(Proteaceae) | |||
4.川苔草科(Podostemales) | 1.川苔草科(Podostemaceae) | ||
1.小二仙草科(Haloragaceae) | |||
2.古奴科(Gunneraceae) | |||
1.海桑科(Sonneratiaceae) | |||
2.千屈菜科(Lythraceae) | |||
3.管萼科(Penaeaceae) | |||
4.隐翼科(Crypteroniaceae) | |||
5.瑞香科(Thymelaeaceae) | |||
6.菱科(Trapaceae) | |||
7.桃金娘科(Myrtaceae) | |||
8.石榴科(Punicaceae) | |||
9.柳叶菜科(Onagraceae) | |||
11.野牡丹科(Melastomataceae) | |||
12.使君子科(Combretaceae) | |||
13.双翼果科(Alzateaceae) | |||
14.谷木科(Memecylaceae) | |||
7.红树目(Rhizophorales) | 1.红树科(Rhizophoraceae) | ||
1.八角枫科(Alangiaceae) | |||
2.紫树科(Nyssaceae) | |||
3.山茱萸科(Cornaceae) | |||
4.绞木科(Garryaceae) | |||
1.毛丝花科(Medusandraceae) | |||
3.铁青树科(Olacaceae) | |||
4.山柚子科(Opiliaceae) | |||
5.檀香科(Santalaceae) | |||
6.羽毛果科(Misodendraceae) | |||
7.桑寄生科(Loranthaceae) | |||
9.房底珠科(Eremolepidaceae) | |||
10.蛇菰科(Balanophoraceae) | |||
1.菌花科(Hydnoraceae) | |||
2.帽蕊草科(Mitrastemonaceae) | |||
3.大花草科(Rafflesiaceae) | |||
11.卫矛目(Celastrales) | 1.四棱果科(Geissolomataceae) | ||
2.卫矛科(Celastraceae) | |||
3.翅子藤科(Hippocrateaceae) | |||
4.木根草科(Stackhousiaceae) | |||
5.刺茉莉科(Salvadoraceae) | |||
6.冬青科(Aquifoliaceae) | |||
7.茶茱萸科(Icacinaceae) | |||
8.鳞枝树科(Aextoxicaceae) | |||
9.心翼果科(Cardiopteridaceae) | |||
10.棒果木科(Corynocarpaceae) | |||
11.毒鼠子科(Dichapetalaceae) | |||
12.苦皮树科(Tepuianthaceae) | |||
1.黄杨科(Buxaceae) | |||
2.油蜡树科(Simmondsiaceae) | |||
3.攀打科(Pandaceae) | |||
4.大戟科(Euphorbiaceae) | |||
1.鼠李科(Rhamnaceae) | |||
2.火筒树科(Leeaceae) | |||
3.葡萄科(Vitaceae) | |||
1.古柯科(Erythroxylaceae) | |||
2.香膏科(Humiriaceae) | |||
3.粘木科(Ixonanthaceae) | |||
4.亚麻藤科(Hugoniaceae) | |||
5.亚麻科(Linaceae) | |||
1.金虎尾科(Malpighiaceae) | |||
2.蜡烛树科(Vochysiaceae) | |||
3.三角果科(Trigoniaceae) | |||
4.孔药花科(Tremandraceae) | |||
5.远志科(Polygalaceae) | |||
6.黄叶树科(Xanthophyllaceae) | |||
7.刺球果科(Krameriaceae) | |||
1.省沽油科(Staphyleaceae) | |||
2.蜜花科(Melianthaceae) | |||
3.钟萼木科(Bretschneideraceae) | |||
4.叠珠树科(Akaniaceae) | |||
5.无患子科(Sapindaceae) | |||
6.七叶树科(Hippocastanaceae) | |||
7.槭树科(Aceraceae) | |||
8.橄榄科(Burseraceae) | |||
9.漆树科(Anacardiaceae) | |||
10.三柱草科(Julianiaceae) | |||
11.苦木科(Simaroubaceae) | |||
13.楝科(Meliaceae) | |||
14.芸香科(Rutaceae) | |||
15.蒺藜科(Zygophyllaceae) | |||
17.牻牛儿苗目(Geraniales) | 1.酢浆草科(Oxalidaceae) | ||
2.牻牛儿苗科(Geraniaceae) | |||
3.池花科(Limnanthaceae) | |||
4.旱金莲科(Tropaeolaceae) | |||
1.五加科(Araliaceae) | |||
2.伞形科(Apiaceae) | |||
6.菊亚纲(Asteridae) | 1.马钱科(Loganiaceae) | ||
2.轮叶科(Retziaceae) | |||
3.龙胆科(Gentianaceae) | |||
4.囊叶木科(Saccifoliaceae) | |||
6.萝藦科(Asclepiadaceae) | |||
1.核果木科(Duckeodendraceae) | |||
2.假茄科(Nolanaceae) | |||
3.茄科(Solanaceae) | |||
4.旋花科(Convolvulaceae) | |||
5.菟丝子科(Cuscutaceae) | |||
6.睡菜科(Menyanthaceae) | |||
7.花葱科(Polemoniaceae) | |||
8.田基麻科(Hydrophyllaceae) | |||
9.异轮叶科(Retziaceae) | |||
1.盖裂寄生科(Lennoaceae) | |||
2.紫草科(Boraginaceae) | |||
3.马鞭草科(Verbenaceae) | |||
1.杉叶藻科(Hippuridaceae) | |||
2.水马齿科(Callitrichaceae) | |||
3.水穗草科(Hydrostachyaceae) | |||
1.车前科(Plantaginaceae) | |||
1.醉鱼草科(Buddlejaceae) | |||
2.木犀科(Oleaceae) | |||
3.玄参科(Scrophulariaceae) | |||
4.球花科(Globulariaceae) | |||
5.苦槛蓝科(Myoporaceae) | |||
6.列当科(Orobanchaceae) | |||
7.苦苣苔科(Gesneriaceae) | |||
8.爵床科(Acanthaceae) | |||
9.胡麻科(Pedaliaceae) | |||
10.紫葳科(Bignoniaceae) | |||
11.对叶藤科(Mendonciaceae) | |||
12.狸藻科(Lentibulariaceae) | |||
|
(Campanulales) | 1.五膜草科(Pentaphragmataceae) | ||
2.楔瓣花科(Sphenocleaceae) | |||
3.桔梗科(Campanulaceae) | |||
4.花柱草科(Stylidiaceae) | |||
5.陀螺果科(Donatiaceae) | |||
6.蓝针花科(Brunoniaceae) | |||
7.草海桐科(Goodeniaceae) | |||
1.茜草科(Rubiaceae) | |||
2.假牛繁缕科(Theligonaceae) | |||
1.忍冬科(Caprifoliaceae) | |||
2.五福花科(Adoxaceae) | |||
3.败酱科(Valerianaceae) | |||
4.川续断科(Dipsacaceae) | |||
1.头花草科(Calyceraceae) | |||
11.菊目(Asterales) | 1.菊科(Asteraceae) | ||
2.Liliopsida(百合纲) | 1.泽泻亚纲(Alismatidae) | 1.花蔺科(Butomaceae) | |
2.沼鳖科(Limnocharitaceae) | |||
3.泽泻科(Alismataceae) | |||
1.水鳖科(Hydrocharitaceae) | |||
3.茨藻目(Najadales) | 1.水蕹科(Aponogetonaceae) | ||
2.冰沼草科(Scheuchzeriaceae) | |||
3.水麦冬科(Juncaginaceae) | |||
4.眼子菜科(Potamogetonaceae) | |||
5.流苏菜科(Ruppiaceae) | |||
6.茨藻科(Najadaceae) | |||
7.角果藻科(Zannichelliaceae) | |||
8.海王草科(Posidoniaceae) | |||
9.丝粉藻科(Cymodoceaceae) | |||
10.大叶藻科(Zosteraceae) | |||
4.霉草目(Triuridales) | 1.无叶莲科(Petrosaviaceae) | ||
2.霉草科(Triuridaceae) | |||
2.槟榔亚纲(Arecidae) | 1.槟榔科(Arecaceae) | ||
1.环花草科(Cyclanthaceae) | |||
1.露兜树科(Pandanaceae) | |||
1.菖蒲科(Acoraceae) | |||
2.天南星科(Araceae) | |||
3.浮萍科(Lemnaceae) | |||
3.鸭拓草亚纲(Commelinidae) | 1.偏穗草科(Rapateaceae) | ||
2.黄眼草科(Xyridaceae) | |||
3.花水藓科(Mayacaceae) | |||
4.鸭跖草科(Commelinaceae) | |||
3.帚灯草目(Restionales) | 1.须叶藤科(Flagellariaceae) | ||
2.假芦苇科(Joinvilleaceae) | |||
3.帚灯草科(Restionaceae) | |||
4.刺鳞草科(Centrolepidaceae) | |||
1.灯心草科(Juncaceae) | |||
2.梭子草科(Thurniaceae) | |||
1.莎草科(Cyperaceae) | |||
2.禾本科(Poaceae) | |||
6.独芯草目(Hydatellales) | 1.独蕊草科(Hydatellaceae) | ||
1.黑三棱科(Sparganiaceae) | |||
2.香蒲科(Typhaceae) | |||
4.姜亚纲(Zingiberidae) | 1.凤梨科(Bromeliaceae) | ||
2.姜目(Zingiberales) | 1.旅人蕉科(Strelitziaceae) | ||
2.赫蕉科(Heliconiaceae) | |||
3.芭蕉科(Musaceae) | |||
4.兰花蕉科(Lowiaceae) | |||
5.姜科(Zingiberaceae) | |||
6.闭鞘姜科(Costaceae) | |||
7.美人蕉科(Cannaceae) | |||
8.竹芋科(Marantaceae) | |||
5.百合亚纲(Liliidae) | 1.田葱科(Philydraceae) | ||
2.雨久花科(Pontederiaceae) | |||
3.血皮草科(Haemodoraceae) | |||
4.蓝星科(Cyanastraceae) | |||
5.百合科(Liliaceae) | |||
6.鸢尾科(Iridaceae) | |||
7.翡若翠科(Velloziaceae) | |||
8.芦荟科(Aloeaceae) | |||
10.刺叶树科(Xanthorrhoeaceae) | |||
11.匍茎草科(Hanguanaceae) | |||
12.箭根薯科(Taccaceae) | |||
13.百部科(Stemonaceae) | |||
14.菝葜科(Smilacaceae) | |||
15.薯蓣科(Dioscoreaceae) | |||
2.兰目(Orchidales) | 1.地鸢尾科(Geosiridaceae) | ||
2.水玉簪科(Burmanniaceae) | |||
3.白玉簪科(Corsiaceae) | |||
4.兰科(Orchidaceae) |
但后来的植物基因遗传学分类研究(被子植物APG 分类法)逐渐让植物学家认知,单子叶植物实际是由古代的双子叶植物演化而来,单子叶植物是双子叶植物的其中一个特化分支,这使传统的双子叶植物分类成为一个并系群而不再被视为有效分类。
主要价值
观赏价值
各地的植物园引种了大量非国产植物(如:中国科学院华南植物园、中国科学院西双版纳热带植物园、上海辰山植物园、深圳仙湖植物园、中国科学院武汉植物园等),据不完全统计己达13,565种,尚不包含大量未鉴定及未列入植物园名录的种。台湾的辜严倬云植物保种中心引种的植物更是多达26,024种。英国皇家园艺学会园艺数据库收录的非中国产商业化栽培植物有48,783种(不包括品种),其中很多种类在国内苗圃、园艺公司都有引种。中国民间植物爱好者也引种了大量的外来植物(薛聪贤,2002-2004;汪远,个人交流)。在仙人掌和多肉植物爱好者的引介下,景天科、番杏科、仙人掌科3个科80%的物种在国内均己有引种,合计约5,000种,再加上其他科的多肉植物,总计达约8,000种。由于兰花爱好者的引介,兰科植物在民间流入中国的外来植物中数量居第一位,估计在10,000种以上。宿根和球根花卉爱好者、水族饲养爱好者等也引入了大量非国产种。电子商务的发展,如淘宝网,更助长了民间引入的力度。据本文作者之一汪远估计,通过各种途径新引入的非国产植物可能多达30,000种,其中FRPS和FOC未记录的属约有1,300个(小属概念),而且这个数字将会持续增长。由于引入的来源、时间、途径不同,所记载的科属归属差别较大,很多外来植物的名称相当混乱。由此可见,为适应学术和社会生产发展的需要,厘清中国原产和引入被子植物的科属名单是极为迫切的。
经济价值
人类的大部分食物和营养来源于被子植物,不是直接地通过农作物或园艺作物如谷类、豆类、薯类、瓜果和蔬菜等,就是间接地为牧场提供牲畜所需的饲料。被子植物还提供建筑、造纸、纺织和塑料制品、油料、纤维、食糖、香料、虫蜡、医药、树脂、鞣酸、麻醉剂、饮料等多得不可计数的原材料。此外,世界上大约有25亿人口从木材和煤炭来获得能源。
被子植物由于它们在种和个体数量上的优势而在覆盖陆地的植物组成中起着主要的作用,形成了作为自然环境景观的大部分植被,并且提供了为大多数陆生动物的生存所需的生境。在生态学上,被子植物对人类的重要意义在于营造防风林和栽种覆盖植物以防止水土流失和海水侵蚀陆地。但人类不仅仅利用自然所赋予的植物,而且还通过植物栽培来改善他们的生活环境,而被子植物在这方面提供了建筑庭园、公园、运动场地、街道绿化、住宅装饰等娱乐和消遣所需的大部分材料。
被子植物在发展人类的文化意识方面起着重要的作用,在宗教和礼仪上占有重要的地位,而且为全世界各族人民的文化提供了大量的象征。茎、叶和花的形状和样式曾被广泛地用作世界上许多地方的艺术、建筑和图案设计的原型。
据估计在农业、林业、果树和蔬菜栽培和生药学上有用的种最少在6000个以上。其中有些在世界贸易中有用,有些则严格说来具有地方性的价值,如那些纯粹属于生产地方性的食物、药物和其他与民间日常生活有关者。此外,还有比这多一倍的种是纯粹的观赏园艺植物。
物种保护
濒危原因
为生境退化或丧失、人为过度利用、物种内在原因、外来入侵种和其他种间竞争、环境污染、自然灾害和极端气候变化等几种类型。这些因子将带来物种居群衰退、分布范围缩小或个体数量下降等不利影响, 从而导致物种受威胁、濒危甚至灭绝。
灭绝原因
被子植物灭绝物种有40种(表2), 分属28科37属。其中特有种29种, 木本植物24种。灭绝种类最多的科依次为兰科(5种)和杜鹃花科(3种); 具有2种的有苦苣苔科、水鳖科、唇形科、樟科、百合科和马鞭草科; 其余20科各含1种。
物种灭绝的主要原因是生境的丧失和退化。人类经济活动改变了土地使用性质, 使野生植物分布地转变为农林植物栽培、养殖和放牧、城镇建设及路网管线建设用地, 从而蚕食毁坏了植物的原生境,造成物种居群数量减少直至消失。还有少数物种的灭绝主要是由于人为过度采集所导致的, 如五加科的三七(Panax notoginseng)和木兰科的绒毛含笑。
|
芒苞草科Acanthochlamydaceae 芒苞草Acanthochlamys bracteata Ⅱ 槭树科Aceraceae 梓叶槭Acer catalpifolium Ⅱ 羊角槭Acer yangjuechi Ⅱ 云南金钱槭Dipteronia dyerana Ⅱ 泽泻科Alismataceae 富宁藤Parepigynum funingense Ⅱ 蛇根木Rauvolfia serpentina Ⅱ 萝藦科Asclepiadaceae 驼峰藤Merrillanthus hainanensis Ⅱ 桦木科Betulaceae 盐桦Betula halophila Ⅱ 金平桦Betula jinpingensis Ⅱ 伯乐树科Bretschneideraceae 伯乐树(钟萼木) Bretschneidera sinensis Ⅰ 花蔺科Butomaceae *拟花蔺Butomopsis latifolia Ⅱ 忍冬科Caprifoliaceae 七子花Heptacodium miconioides Ⅱ 石竹科Caryophyllaceae 卫矛科Celastraceae 膝柄木Bhesa sinensis Ⅰ 永瓣藤Monimopetalum chinense Ⅱ 连香树科Cercidiphyllaceae 连香树Cercidiphyllum japonicum Ⅱ 使君子科Combretaceae 萼翅藤Calycopteris floribunda Ⅰ 千果榄仁Terminalia myriocarpa Ⅱ 菊科Compositae *画笔菊Ajaniopsis penicilliformis Ⅱ *革苞菊Tugarinovia mongolica Ⅰ 四数木科Datiscaceae 四数木Tetrameles nudiflora Ⅱ 龙脑香科Dipterocarpaceae 东京龙脑香Dipterocarpus retusus Ⅰ 狭叶坡垒Hopea chinensis Ⅰ 无翼坡垒(铁凌) Hopea exalata Ⅱ 坡垒Hopea hainanensis Ⅰ 多毛坡垒Hopea mollissima Ⅰ 望天树Parashorea chinensis Ⅰ 广西青梅Vatica guangxiensis Ⅱ 茅膏菜科Droseraceae *貉藻Aldrovanda vesiculosa Ⅰ 胡颓子科Elaeagnaceae 翅果油树Elaeagnus mollis Ⅱ 大戟科Euphorbiaceae 东京桐Deutzianthus tonkinensis Ⅱ 壳斗科Fagaceae 华南锥Castanopsis concinna Ⅱ 台湾水青冈Fagus hayatae Ⅱ 三棱栎Formanodendron doichangensis Ⅱ 瓣鳞花科 Frankeniaceae *瓣鳞花Frankenia pulverulenta Ⅱ 龙胆科Gentianaceae *辐花Lomatogoniopsis alpina Ⅱ 苦苣苔科Gesneriaceae 瑶山苣苔Dayaoshania cotinifolia Ⅰ 单座苣苔Metabriggsia ovalifolia Ⅰ 秦岭石蝴蝶Petrocosmea qinlingensis Ⅱ 报春苣苔Primulina tabacum Ⅰ 辐花苣苔Thamnocharis esquirolii Ⅰ 禾本科Gramineae 酸竹Acidosasa chinensis Ⅱ *沙芦草Agropyron mongolicum Ⅱ *异颖草Anisachne gracilis Ⅱ *短芒披碱草Elymus breviaristatus Ⅱ *无芒披碱草Elymus submuticus Ⅱ *毛披碱草Elymus villifer Ⅱ *内蒙古大麦Hordeum innermongolicum Ⅱ *药用野生稻Oryza officinalis Ⅱ *普通野生稻Oryza rufipogon Ⅱ *四川狼尾草Pennisetum sichuanense Ⅱ *华山新麦草Psathyrostachys huashanica Ⅰ *三蕊草Sinochasea trigyna Ⅱ *拟高粱Sorghum propinquum Ⅱ *箭叶大油芒Spodiopogon sagittifolius Ⅱ *中华结缕草Zoysia sinica Ⅱ 小二仙草科Haloragidaceae *乌苏里狐尾藻Myriophyllum ussuriense Ⅱ 山铜材Chunia bucklandioides Ⅱ 长柄双花木Disanthus cercidifolius var. longipes Ⅱ 半枫荷Semiliquidambar cathayensis Ⅱ 银缕梅Shaniodendron subaequalum Ⅰ 四药门花Tetrathyrium subcordatum Ⅱ 水鳖科Hydrocharitaceae *水菜花Ottelia cordata Ⅱ 子宫草Skapanthus oreophilus Ⅱ 樟科Lauraceae 油丹Alseodaphne hainanensis Ⅱ 普陀樟Cinnamomum japonicum Ⅱ 油樟Cinnamomum longepaniculatum Ⅱ 卵叶桂Cinnamomum rigidissimum Ⅱ 润楠Machilus nanmu Ⅱ 闽楠Phoebe bournei Ⅱ 浙江楠Phoebe chekiangensis Ⅱ 楠木Phoebe zhennan Ⅱ 豆科Leguminosae *线苞两型豆Amphicarpaea linearis Ⅱ 黑黄檀(版纳黑檀) Dalbergia fusca Ⅱ 格木Erythrophleum fordii Ⅱ 山豆根(胡豆莲) Euchresta japonica Ⅱ 绒毛皂荚Gleditsia japonica var. velutina Ⅱ *野大豆Glycine soja Ⅱ *烟豆Glycine tabacina Ⅱ *短绒野大豆Glycine tomentella Ⅱ 红豆树Ormosia hosiei Ⅱ 缘毛红豆Ormosia howii Ⅱ 紫檀(青龙木) Pterocarpus indicus Ⅱ 油楠(蚌壳树) Sindora glabra Ⅱ 任豆(任木) Zenia insignis Ⅱ 狸藻科Lentibulariaceae *盾鳞狸藻Utricularia punctata Ⅱ 木兰科Magnoliaceae 长蕊木兰Alcimandra cathcardii Ⅰ 地枫皮Illicium difengpi Ⅱ | 单性木兰Kmeria septentrionalis Ⅰ 大叶木兰Magnolia henryi Ⅱ 馨香玉兰Magnolia odoratissima Ⅱ 厚朴Magnolia officinalis Ⅱ 凹叶厚朴Magnolia officinalis subsp. biloba Ⅱ 长喙厚朴Magnolia rostrata Ⅱ 西康玉兰Magnolia wilsonii Ⅱ 宝华玉兰Magnolia zenii Ⅱ 香木莲Manglietia aromatica Ⅱ 大果木莲Manglietia grandis Ⅱ 毛果木莲Manglietia hebecarpa Ⅱ 华盖木Manglietiastrum sinicum Ⅰ 石碌含笑Michelia shiluensis Ⅱ 峨眉含笑Michelia wilsonii Ⅱ 峨眉拟单性木兰Parakmeria omeiensis Ⅰ 云南拟单性木兰Parakmeria yunnanensis Ⅱ 合果木Paramichelia baillonii Ⅱ 水青树Tetracentron sinense Ⅱ 楝科Meliaceae 粗枝崖摩Amoora dasyclada Ⅱ 红椿Toona ciliata Ⅱ 毛红椿Toona ciliata var. pubescens Ⅱ 防己科Menispermaceae 藤枣Eleutharrhena macrocarpa Ⅰ 肉豆蔻科Myristicaceae 海南风吹楠Horsfieldia hainanensis Ⅱ 滇南风吹楠Horsfieldia tetratepala Ⅱ 茨藻科Najadaceae *高雄茨藻Najas browniana Ⅱ *拟纤维茨藻Najas pseudogracillima Ⅱ 睡莲科Nymphaeaceae *莼菜Brasenia schreberi Ⅰ *莲Nelumbo nucifera Ⅱ *贵州萍逢草Nuphar bornetii Ⅱ 蓝果树科Nyssaceae 喜树(旱莲木) Camptotheca acuminata Ⅱ 珙桐Davidia involucrata Ⅰ 光叶珙桐Davidia involucrata var. vilmoriniana Ⅰ 云南蓝果树Nyssa yunnanensis Ⅰ 金莲木科Ochnaceae 合柱金莲木Sinia rhodoleuca Ⅰ 铁青树科Olacaceae 蒜头果Malania oleifera Ⅱ 木犀科Oleaceae 水曲柳Fraxinus mandshurica Ⅱ 棕榈科Palmae 董棕Caryota urens Ⅱ 小钩叶藤Plectocomia microstachys Ⅱ 龙棕Trachycarpus nana Ⅱ 罂粟科Papaveraceae 斜翼科Plagiopteraceae 斜翼Plagiopteron suaveolens Ⅱ 川苔草科Podostemaceae *川藻(石蔓) Terniopsis sessilis Ⅱ 蓼科Polygonaceae 毛茛科Ranunculaceae 粉背叶人字果Dichocarpum hypoglaucum Ⅱ 独叶草Kingdonia uniflora Ⅰ 马尾树科Rhoipteleaceae 马尾树Rhoiptelea chiliantha Ⅱ 茜草科Rubiaceae 绣球茜Dunnia sinensis Ⅱ 香果树Emmenopterys henryi Ⅱ 丁茜Trailliaedoxa gracilis Ⅱ 芸香科Rutaceae 黄檗(黄菠椤) Phellodendron amurense Ⅱ 川黄檗(黄皮树) Phellodendron chinense Ⅱ 杨柳科Salicaceae 钻天柳Chosenia arbutifolia Ⅱ 无患子科Sapindaceae 伞花木Eurycorymbus cavaleriei Ⅱ 掌叶木Handeliodendron bodinieri Ⅰ 山榄科Sapotaceae 紫荆木Madhuca pasquieri Ⅱ 蛛网萼Platycrater arguta Ⅱ 冰沼草科Scheuchzeriaceae *冰沼草Scheuchzeria palustris Ⅱ 玄参科Scrophulariaceae *胡黄连Neopicrorhiza scrophulariiflora Ⅱ 呆白菜(崖白菜) Triaenophora rupestris Ⅱ 茄科Solanaceae *山莨菪Anisodus tanguticus Ⅱ 黑三棱科Sparganiaceae *北方黑三棱Sparganium hyperboreum Ⅱ 梧桐科Sterculiaceae 广西火桐Erythropsis kwangsiensis Ⅱ 蝴蝶树Heritiera parvifolia Ⅱ 平当树Paradombeya sinensis Ⅱ 景东翅子树Pterospermum kingtungense Ⅱ 猛仑翅子树Pterospermum menglunense Ⅱ 安息香科Styracaceae 长果安息香Changiostyrax dolichocarpa Ⅱ 秤锤树Sinojackia xylocarpa Ⅱ 瑞香科Thymelaeaceae 土沉香Aquilaria sinensis Ⅱ 椴树科Tiliaceae 柄翅果Burretiodendron esquirolii Ⅱ 蚬木Burretiodendron hsienmu Ⅱ 滇桐Craigia yunnanensis Ⅱ 海南椴Hainania trichosperma Ⅱ 紫椴Tilia amurensis Ⅱ 菱科Trapaceae *野菱Trapa incisa Ⅱ 榆科Ulmaceae 长序榆Ulmus elongata Ⅱ 榉树Zelkova schneideriana Ⅱ 伞形科Umbelliferae *珊瑚菜(北沙参) Glehnia littoralis Ⅱ 马鞭草科Verbenaceae 海南石梓(苦梓) Gmelina hainanensis Ⅱ 姜科Zingiberaceae 茴香砂仁Etlingera yunnanense Ⅱ 拟豆蔻Paramomum petaloideum Ⅱ 长果姜Siliquamomum tonkinense Ⅱ |
保护级别
被子植物也是我国濒危植物的主体, 许多种类已经被列入国内保护植物名录和国际公约保护物种名录。 我国共有约1,600种植物被列入, 其中绝大部分是被子植物的兰科植物。
植物文化
花图式
为了简单地说明一朵花的结构,花各部分的组成、排列位置和相互关系,可以用一个公式或图案把一朵花的各部分表示出来,前者称为花程式(floral formula),后者称为花图式(floral diagram)。花图式(flower diagram)实际上就是花的各部分在垂直于花轴的平面上的投影,并用各部分横切面简图表示其实际情况所得到的图形。
花图式是花的横切面简图,用以表示花各部分的轮数、数目、离合、排列(包括花被卷叠方式)、胎座式等。花图式还能表示花的远轴面和近轴面,在绘制花图式时,用黑色圆点在花图式上方表示花着生的花轴;用空心的弧片在花图式的两侧表示苞片;用带有线条的弧片表示萼片,弧片中央尖突的部分表示萼片的中脉;实心的弧片表示花瓣,雄蕊和雌蕊就用花药和子房的横切片面来表示;用连接线表示雄蕊的连合或与花冠的贴生;子房的胎座式应绘出子房室数和胚珠的着生方式。
2.雌花器:花的中心位置为心皮(雌花器),常见为空心圆线,模拟子房横切面;圆中可以实线或虚线的分隔线表示心皮数目及黏合状态(合生或离生的心皮、子房室壁的状态),胎座型以及胚珠着生情况等。
3.雄蕊:雄蕊常采用2个空心小圆圈表示花药的横切面,并表示出现相应的排列方式、轮数、合生或离生等;假雄蕊常采同样图形、但为实心。
4.花被:视花被是否区分为花萼及花瓣(或花冠,合生的花瓣),并依萼瓣合生或离生相互连接或分离。
花公式
花公式或称为花程式,是使用符号和数字来表达花的各部分组成、  排列、位置及彼此间的关系。书写顺序是性别、整齐与否、花被(花萼和花冠)、雄蕊群、雌蕊群。各个符号含义如下:K代表花萼(Kelch)
排列、位置及彼此间的关系。书写顺序是性别、整齐与否、花被(花萼和花冠)、雄蕊群、雌蕊群。各个符号含义如下:K代表花萼(Kelch)
C代表花冠(Corolla)
A代表雄蕊群(Androeciurn)
G代表雌蕊群(Gynoecium)
P代表花被(Perianth)
花各部分的数目可用数字来表示,如该部分缺少时就用“0”表示,数目很多就用“∞”来表示,并把它们写于代表各部分字母的右下角处。
如果某部分在一轮以上时就用“+”来表示,如果某一部分其个体相互连合就用“( )”表示。
子房的位置可以在雌蕊的字母下边加一道横线表示上位子房( ),在上面加一道横线表示下位子房( ),上下各加一道横钱表示半下位子房( )。
辐射对称花(整齐花)用“*”表示;两侧对称花(不整齐花)用“↑”表示;♀表示雌花,♂表示雄花, 表示两性花,书写在花程式的前边。
相关作品
附件列表
词条内容仅供参考,如果您需要解决具体问题
(尤其在法律、医学等领域),建议您咨询相关领域专业人士。