黄山梅

形态特征

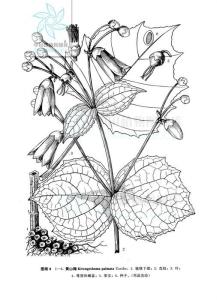
 多年生草本,高80-120米。茎直立,近四棱形,无毛,略紫色。叶生于茎下部的最大,圆心形,长和宽均10-20厘米,掌状7-10裂,裂片具粗齿,基部近心形,两面被糙伏毛,生于茎上部的叶渐较小,长、宽均3-7厘米,最上部的叶卵形或披针形,先端渐尖;叶柄生于茎最下部的最长,长达25厘米,生于茎上部的渐短以至于无柄。聚伞花序生于茎上部叶腋及顶端,通常具3花,中部的花最大,无小苞片,两侧的花较小,具小苞片,有时退化仅具1-2花;苞片披针形;花黄色,直径4-5厘米;花梗长1-3(-4)厘米,中部的花梗常较两侧的短,稍被紧贴柔毛;萼筒半球形,直径7-10毫米,被柔毛,裂片三角形;花瓣5,离生,形状稍不等,长圆状倒卵形或近狭倒卵形,长2.5-3.5厘米,宽10-15毫米,先端急尖;雄蕊15,外轮的与花瓣近等长,内轮的稍短;花柱线形,向上稍狭,长约2厘米。蒴果阔椭圆形或近球形,直径约1.3厘米,顶端具宿存花柱,干时褐色;种子黄色,扁平,连翅长7-10毫米,宽3-5毫米,周围具膜质斜翅。花期3-4月,果期5-8月。
多年生草本,高80-120米。茎直立,近四棱形,无毛,略紫色。叶生于茎下部的最大,圆心形,长和宽均10-20厘米,掌状7-10裂,裂片具粗齿,基部近心形,两面被糙伏毛,生于茎上部的叶渐较小,长、宽均3-7厘米,最上部的叶卵形或披针形,先端渐尖;叶柄生于茎最下部的最长,长达25厘米,生于茎上部的渐短以至于无柄。聚伞花序生于茎上部叶腋及顶端,通常具3花,中部的花最大,无小苞片,两侧的花较小,具小苞片,有时退化仅具1-2花;苞片披针形;花黄色,直径4-5厘米;花梗长1-3(-4)厘米,中部的花梗常较两侧的短,稍被紧贴柔毛;萼筒半球形,直径7-10毫米,被柔毛,裂片三角形;花瓣5,离生,形状稍不等,长圆状倒卵形或近狭倒卵形,长2.5-3.5厘米,宽10-15毫米,先端急尖;雄蕊15,外轮的与花瓣近等长,内轮的稍短;花柱线形,向上稍狭,长约2厘米。蒴果阔椭圆形或近球形,直径约1.3厘米,顶端具宿存花柱,干时褐色;种子黄色,扁平,连翅长7-10毫米,宽3-5毫米,周围具膜质斜翅。花期3-4月,果期5-8月。
 |  |
生长特性
黄山梅为 阴性草木,不耐强光照射,喜温凉、湿润、富含有机质的酸性黄棕壤的生境,常在落叶阔叶林下阴湿之地呈小片生长。分布区年平均温约7.7℃,1月平均温-3.4℃,7月平均温17.8℃,年降水量约2000毫米,相对湿度约90%。浙江西部其上层树种主要为灯台树Cornus controversa Hemsl.、华东桤叶树Clethra barbinervis Sieb et Zucc.、圆锥绣球Hydrangea paniculata Sieb. 等;伴生植物主要有窄头橐吾Ligularia stenocephala (Maxim.) Matsum. et Koidz.、油点草Tricyrtis macropoda Miq.与阔鳞鳞毛蕨Dryopteris championii (Benth.) C.Chr. ex Ching等。本种新芽于3月上中旬萌动出土,4月上旬展叶。花蕾于6月下旬出现,7月上旬开花,可延至8月。蒴果于9--11月成熟。
分布范围
产安徽(黄山)和浙江(天目山)。生于海拔700-1800米山谷林中阴湿处。日本、朝鲜亦产。模式标本采自日本。
 黄山梅分布区域较窄,且呈跳动性分布,具有重要的科研价值。在中国主要分布于安徽与黄山和浙江天目山,由于森林砍伐,生境破坏以及挖根入药等原因,致使植株日益减少,已处于濒临状态。此外,日本也有少量分布。黄山梅生长的环境条件比较持殊;在黄山地区大多生长在海拔1500一1700米高山的北坡,如北海的峡谷中就有成片的分布。多与天女花、马褂木,长柄山毛掸、黄山杜鹃、中华、龙须草等伴生。它比较耐寒,能耐零下20℃的低温。对土壤要求比较严格,天然分布区内土壤均为高山腐殖质土,有机质丰富,排水透气性良好。它要求有较高的相对湿度,年平均湿度须达75%以广,并且经常处在云雾缭绕之中。笔者曾做过引种驯化试验,在海拔300一400米的地带,选择遮荫良好的场所种植,也能正常越夏和开花结实。
黄山梅分布区域较窄,且呈跳动性分布,具有重要的科研价值。在中国主要分布于安徽与黄山和浙江天目山,由于森林砍伐,生境破坏以及挖根入药等原因,致使植株日益减少,已处于濒临状态。此外,日本也有少量分布。黄山梅生长的环境条件比较持殊;在黄山地区大多生长在海拔1500一1700米高山的北坡,如北海的峡谷中就有成片的分布。多与天女花、马褂木,长柄山毛掸、黄山杜鹃、中华、龙须草等伴生。它比较耐寒,能耐零下20℃的低温。对土壤要求比较严格,天然分布区内土壤均为高山腐殖质土,有机质丰富,排水透气性良好。它要求有较高的相对湿度,年平均湿度须达75%以广,并且经常处在云雾缭绕之中。笔者曾做过引种驯化试验,在海拔300一400米的地带,选择遮荫良好的场所种植,也能正常越夏和开花结实。
 |  |
发现来源
黄山梅是多年生草本植物,最早发现于 日本。1888年8月9日日本 植物分类学家矢天部良吉在伊予国爱媛县石锤山首次采集到开花期的黄山梅标本;1890年10月吉永虎马又在同一地方采集到果实期的黄山梅标本, 模式标本。1890年,矢天部良吉在日本刊物《植物学杂志》上以 英文形式正式首次发表黄山梅这一新属新种,定名为Kirengeshoma palmata Yatabe。人们最初认为黄山梅是日本四国地区的特产,不过后来在纪伊半岛和九州也相继 发现。1935年,日本 东京大学植物分类学教授中井猛之进在 朝鲜半岛全罗南道的白羊山又采集到黄山梅,记载为Kirengeshoma koreana Nakai。另外,1978年鹿儿岛大学教授初岛住彦,按照远藤正喜在大分县祖母山上游溪谷(大野郡绪方町尾平)采集的标本,又发表了Kirengeshoma suffruticosaHatusima。
但是,后来经过研究发现,上述两植物都是黄山梅种下的变异类型,因而与 黄山梅归并,成为黄山梅的异名。除日本和韩国外,黄山梅还分布在中国安徽省黄山地区包括黄山风景区及 歙县清凉峰,和 浙江省天目山地区,即浙江天目山和 昌化龙塘山。
生存环境
黄山梅为多年生草本植物,该植物仅间断分布于 中国和日本,为典型的中国-日本特有种。在日本,该植物主要分布于日本的中西部,包括本州的纪伊半岛、九州、四国等地,南韩半岛也有分布;在 中国,黄山梅只斑块状分布于华东地区的皖浙交界处,具体包括:皖南山区的安徽黄山自然风景区(HS)、歙县清凉峰自然保护区(QLF)和浙西北地区的天目山自然保护区(TMS)、昌化龙塘山自然保护区(LTS)。植株多零星或集群分布,海拔高度范围为700m-1500m,多生长于林下阴湿处、路边或花岗岩岩石上黄山梅大都生长在遮荫度较高的林下石壁或流水的石缝处。与之伴生的灌木或小乔木植物有 天女花、圆锥绣球、 金缕梅、 白檀、 黄山杜鹃、鸡桑及 小构树等;草本层主要为 小升麻、 三脉紫菀、华南落新妇、 细辛、 及己、 唐松草、 凹叶景天、 车前、 油点草以及 六叶葎等,另外还有成片的 苔藓及 蕨类植物。其中天女花、圆锥绣球、小升麻、凹叶景天、苔草以及苔藓植物是其群落组成的主要成分。
黄山梅分布区郁闭度较高,达到80%-90%(夏季),地表潮湿,常有苔藓和 景天科等喜湿性植物覆盖,土壤肥沃,腐殖质明显。对生境的严格要求可能是限制其自然分布的一个重要因素。
种群分布格局是物种与环境长期以来相互适应、相互作用的结果,它不仅与物种的生物学特性和种群间的竞争排斥有关,而且与物种的生境(包括土壤、地形、地貌等)有密切关系。黄山梅种群的分布格局与大多数珍稀濒危植物自然种群一样,呈现聚集分布式样。其主要原因是:首先,黄山梅为多年生草本植物,可以利用根状茎进行 无性繁殖。在无性繁殖中,黄山梅的无性系 克隆植株的产生是依靠其根状茎断裂脱离母体而形成,克隆植株集生于母株周围,导致其聚集程度较高。其次,对环境条件的严格要求是导致黄山梅聚集分布的另一原因。黄山梅仅分布于皖浙交界处的部分山区,即安徽地区的黄山风景区、歙县清凉峰,浙江地区的天目山和昌化龙塘山,海拔高度范围为700m-1500m,阴坡及半阴坡的林下,伴生植物主要有圆锥绣球、天女花等,群落的郁闭度多在80%-90%,林下阴暗潮湿,光照较为微弱,很少受到阳光的直接照射,空气和 土壤的 相对湿度较大。尤其在林木遮荫下的大块 岩石小生境或有藓类植物和景天科植物覆盖、土壤腐殖质较厚的地段黄山梅生长良好。另外,生境的异质性限制了黄山梅的分布,导致其聚集生长。黄山梅生长在中高海拔的林下山地的花岗岩地带,土层较薄,小范围内生境的差异性较大,岩石的阻碍也使根状茎的延伸受到限制。
繁殖过程
黄山梅为多年生草本植物。十二月中下旬,地下茎进入休眠状态,在地下茎结节处形成一枚由多层鳞片包裹的越 冬芽,芽不露出土面。三月上中旬新芽由地下根状茎萌动,通过其基部的居间生长将整个芽推出地面,嫩芽锥形,粉紫色。四月上旬开始展叶,随后进行快速的营养生长。六月下旬出现 花蕾, 茎顶端的花蕾首先出现。此时,营养生长基本停止,开始进入 生殖器官的发育阶段。七月上旬开始偶见花开,中旬为盛花期,有昆虫来访。发育较迟的花可延迟至八月中下旬。但直至花瓣即将脱落,花瓣都不完全张开。九月座果,十月中下旬至十一月 蒴果成熟,种子由蒴果背缝线开裂处散出。十一月下旬,黄山梅地上茎开始枯萎死亡。据野外观察发现,黄山梅的传粉昆虫主要为 蜂类和 蝇类。另外发现这些昆虫的专一性较差,在访问黄山梅的同时,还拜访 菊科、 唇形科及虎耳草科等其它伴生植物,如三脉紫菀、华南落新妇等。访花昆虫的高峰期通常在上午8:00-10:00和下午15:00-17:00左右,阴天及小雨天也能见其频繁活动,而正午阳光照射最强烈时很少看到其踪影。一般情况下,花冠张开2个 小时左右,花粉基本上就可全部散出,风力也在一定程度上促进了花粉的传播。
种子结构
黄山梅种子结构很特殊:外被由单层、排列致密的细胞构成的网格状外种皮,即种翅;内种皮与较为发达的 胚乳紧密结合,细胞排列紧密,且显著木质化增厚;中央包被着成熟的双 子叶胚。由于黄山梅的胚埋于胚乳之中,保护组织坚硬,机械性强,且外罩一层膜质外种皮,这对抵御严寒和病原菌的侵蚀来说是有利的,但同时这一结构又对种子萌发较为不利。机械组织的阻挡,尤其是不易获取 氧气和 水分以及营养物质而限制了种子的萌发,这可能是造成黄山梅种子在自然条件下萌发率低、实生苗少的原因之一。
保护状况
保护价值
黄山梅为单种属植物,是黄山梅亚科Kirengeshomoideae唯一的代表种,也是中国、日本间断分布的典型种类。对于阐明虎耳草科的种系演化以及中国和日本植物区系的关系,均有科研价值。
保护措施
栽培要点
黄山梅的繁殖和栽培并不十分困难。可采用播种育苗,但 种子发芽率不很高,且生长缓慢,通常情况下可采用分株繁殖,深秋或早春,从野外连根掘起块茎,再从块茎间结合比较薄弱处摊开,每一块都须具有1—2芽,然后进行栽种,当然,要注重保护资源,切忌乱掘滥挖。栽培用土可选高山腐殖土,浇灌用水以河水和糖水为好,切忌水中含有氯离子及 碱性物质,否则会造成烂根。为减少夏天遮荫的麻烦,可将植株栽种在大树的间隙中,借树冠的浓荫,降低地面的温度和减小光照强度。炎热和夏季,植株叶面要经常喷水,以确保其安全越夏。
特征研究
对黄山梅属(Kirengeshoma Yatabe)黄山梅(Kirengeshoma palmata Yatabe)及其相关植物--草绣球属(Cardiandra Sieb.&Zucc.)和银梅草属(Deinanthe Maxim.)植物作了叶表皮的微形态研究.结果表明:黄山梅(Kirengeshoma palmata Yatabe)叶下表皮细胞不规则,垂周壁深波状,气孔类型为不规则型;叶上表皮不规则或近多角形,垂周壁平直、弓形或波状.叶上、下表面均具单细胞T形毛,毛上疣突显著。毛基为单细胞型.草绣球(Cardiandra mollendorffii Li)叶下表皮不规则,垂周壁深波状,气孔类型为不规则型;叶上表皮细胞不规则,垂周壁浅波状,具针晶,单毛,具鞘状结构,表面光滑。毛基为多细胞型。二裂叶银梅草(Deinanthe bifida Maxim.)和银梅草(Deinanthe caerulea Stapf.)叶下表皮细胞不规则,垂周壁深波状,气孔类型为不规则型;叶上表皮细胞形状与垂周壁式样同叶下表皮,且均具针晶。另外,在二裂叶银梅草叶上、下表面发现两种类型的毛:锥形毛和T形毛。结合三属的现有资料,通过比较分析,笔者初步认为: 三属性状高度重叠,关系甚密,银梅草属可能是黄山梅属和草绣球属的过渡类型;叶表皮特征观察结果不支持Takhtajan系统把黄山梅独立出来提升为亚科的观点。
药用价值

附件列表
词条内容仅供参考,如果您需要解决具体问题
(尤其在法律、医学等领域),建议您咨询相关领域专业人士。